
- Типы питания прокариот. Группы микроорганизмов по типу питания
- Какие группы бактерий по способу питания могут быть и в чем их различия
- Принципиальные отличия питания двух групп микробов
- Автотрофные микроорганизмы
- Гетеротрофные бактерии
- Значение бактерий разного способа питания для природы и человека
- Типы питания прокариот
Типы питания прокариот. Группы микроорганизмов по типу питания
В отличие от растительных и животных организмов, имеющих один вполне определенный тип питания – соответственно автотрофный и гетеротрофный, прокариоты характеризуются многообразием типов питания. Поэтому для характеристики типов питания трокариотных организмов используются одновременно три критерия: источник углерода, источник энергии и донор электронов (водорода).
По источнику углерода прокариоты являются автотрофами, если они получают углерод в результате фиксации углекислого газа, и гетеротрофами, если источником углерода для них служат органические соединения.
По источнику энергии прокариоты, использующие солнечный свет, называются фототрофами, а получающие энергию за счет окислительно-восстановительных реакций – хемотрофами.
И наконец, по донору электронов прокариоты подразделяются на литотрофы, обладающие способностью использовать неорганические вещества, органотрофы, использующие в качестве доноров электрона органические соединения.
По трем вышеуказанным критериям выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы хемолитоавтотрофы и хемоорганогетеротрофы (таблица 1).
Различают следующие типы гетеротрофии: паразитизм облигатный внутриклеточный, паразитизм факультативный, сапрофитизм.
Организмы, которые могут жить только внутри других организмов, имеют редуцированный метаболизм, зависят от метаболизма хозяина, являются облигатными внутриклеточными паразитами.
Факультативные паразиты-организмы, способные расти вне клетки хозяина при подходящих условиях, на питательных средах.
Сапрофиты – гетеротрофные организмы, нуждаются в готовых органических веществах, непосредственно от организмов не зависят.
Сапрофиты нуждаются в разных концентрациях органических веществ. Олиготрофы – способны расти при низких концентрациях органического вещества (1–15 мг углерода в литре раствора). Копиотрофы – предпочитают высокие концентрации питательных веществ (10 грамм углерода в литре раствора).
Дата добавления: 2015-08-14 ; просмотров: 4903 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Какие группы бактерий по способу питания могут быть и в чем их различия
Каждый живой организм, даже одноклеточный, которым является бактерия, нуждается в питательных веществах. Царство микробов огромно, его представителей отличают между собой по многим признакам. В частности, по способу питания бактерии делятся на автотрофные и гетеротрофные организмы.

Микроорганизмы автотрофного и гетеротрофного способов питания – два огромных звена в круговороте веществ на Земле. Первые создают начальные органические элементы для жизнеобеспечения следующего яруса живых организмов.
Вторые формируют биомассу для питания животных и растений, сопровождают их в течение жизни, затем уничтожают мертвые тела вплоть до неорганических веществ, создавая пищу для автотрофов. Цикл начинается снова.
Принципиальные отличия питания двух групп микробов
Разделение бактерий на две группы стало возможным после изучения их обменных процессов. Оказалось, что одни обладают большей самостоятельностью, чем другие.
- Автотрофам свойственно питание неорганическими веществами простейшей структуры (водород, азот, углерод и другие). Из них бактерия самостоятельно создает сложные органические конструкции для жизнедеятельности.
- Гетеротрофы нуждаются в готовых органических элементах для своего питания.
Оба вида организмов могут поглощать необходимые им вещества только в виде растворов, поэтому важнейший элемент бактериального питания – вода. Еще она является поставщиком кислорода и водорода для реакций окисления/восстановления.
Автотрофные микроорганизмы
По способам питания автотрофные бактерии относятся к двум подгруппам в зависимости от своего энергообеспечения:
- фотосинтезирующие бациллы, использующие для обменных процессов энергию светового излучения;
- хемосинтезирующие организмы, которые для достижения этих целей прибегают к окислительно-восстановительным реакциям.
Автотрофы относятся преимущественно к почвенным микроорганизмам. Они обогащают почву органикой, что делает ее более плодородной. Активно «трудятся» в приземном слое, превращая опавшую листву, пожухлую траву в питательный гумус.
Фактически автотрофы создают органические вещества, которые используются в питании гетеротрофными организмами. А неорганические остатки колоний автотрофных железобактерий через тысячи лет могут стать месторождениями одноименных руд.
Человеческий организм для бактерий данного типа питания не представляет интереса в качестве среды обитания. Тела людей состоят из сложных органических соединений. Для автотрофных микробов они «несъедобны». По-другому относятся к человеческому организму как питательной среде бациллы гетеротрофного способа питания.
Гетеротрофные бактерии
Эти представители микромира существуют за счет других живых организмов, употребляя в пищу составляющие их органические вещества. Есть три исхода «встречи» бактерии-гетеротрофа и хозяина:
- микроб убивает животное или растение;
- иммунная защита хозяина уничтожает бациллу;
- возникает неактивное бактерионосительство или взаимовыгодное сосуществование.
Один и тот же микроорганизм в разных условиях может быть участником любой из трех ситуаций. Первоначальное его попадание в сильный молодой организм закончится смертью микроба или бактерионосительством. Как только организм хозяина ослабеет, бацилла активируется и убьет его.
По признакам взаимоотношений бактерий-гетеротрофов с живыми организмами их условно относят к трем большим группам.
- Патогенные микробы, которые, паразитируя в организме жертвы, вызывают у нее инфекционные заболевания.
- Сапрофитная флора – тихий паразит. Эти микробы могут жить в организме хозяина, не причиняя ему неприятностей. Они питаются омертвевшими клетками, остатками веществ, которые прошли через систему пищеварения хозяина.
- Симбиотические микроорганизмы взамен потребляемых ресурсов организма хозяина вырабатывают для него полезные вещества. Например, клубеньковые бактерии растений или витаминопродуцирующие микроорганизмы кишечника человека. Иногда эта взаимозависимость настолько сильна, что в случае гибели микрофлоры умирает ее бывший носитель.
Значение бактерий разного способа питания для природы и человека
Автотрофные бактерии имеют узкую «специализацию», но от этого их значение для природы не становится меньше, чем гетеротрофов. Автотрофы создают основу для всего органического многообразия на нашей планете. Многие из них становятся «создателями» минеральных месторождений.
У гетеротрофных микробов функций больше.
- Естественный отбор, который они осуществляют, уничтожая слабые, больные и старые организмы.
- Помощь в жизнеобеспечении (клубеньковые бактерии у растений, вырабатывающие витамины – у животных).
- Санитарная роль состоит в гнилостном разложении останков живых существ.
Роль бактерий-гетеротрофов в естественном отборе ясна и прозрачна. Природа стремится к совершенству, поэтому слабые организмы уничтожаются, давая место для появления более сильных особей. Устраняются и генетически несовершенные субъекты, возникающие в процессе эволюции.
Оставшимся оказывается помощь для их развития. Показательны два примера из растительной и животной жизни.
- Клубеньковые бактерии служат обогащению почвы азотом из воздуха. Они имеют специальные элементы (мезосомы), которые фиксируют азот из окружающей среды. Живут клубеньковые бактерии в корнях растений семейства бобовых.
Попадают они туда через микротрещины, потом выделяют вещества, которые стимулируют размножение клеток корня. На нем возникают клубеньковые утолщения. В них клубеньковые бактерии накапливают азот для обмена с растением на углеводы.
Этот феномен синергизма клубеньковых растений и микроорганизмов люди используют в сельском хозяйстве. Бедные азотом почвы засеивают бобовыми растениями, клубеньковые бактерии которых обогащают их азотом.
По осени их запахивают в землю. Так необходимый азот попадает из погибших растений и клубеньковых микроорганизмов в почву для последующего употребления другими культурами, которыми засеют это поле.
- Кишечник животных изнутри выстлан гетеротрофными бактериями, которые вырабатывают витамины группы B и K. Таким образом, недостаток их в пище животных и человека восполняют бактерии-симбионты гетеротрофного способа питания.
Помимо этого, гетеротрофы используются для квашения овощей, бродильных процессов. Одним из таких является молочнокислое брожение. В результате получается большое разнообразие молочнокислых продуктов, необходимых для питания человека.
Многие люди имеют аллергию на цельное молоко домашних животных. Употребление молочнокислых продуктов такой реакции не вызывает, потому что белок в них денатурирован. А он ответственен за развитие аллергических реакций.
Заключительная роль гетеротрофов в жизни каждого существа – гнилостное разложение его органических остатков. Процессы гниения необходимы природе так же, как и возникновение жизни. Микроорганизмы, осуществляющие гнилостные разрушения органики, в этот период очень опасны.
Во время гниения даже сапрофиты и симбионты могут переродиться в хищников. Гнилостные массы имеют в своем составе высокие концентрации высокопатогенных микроорганизмов. Но без такой «грязной» работы бактерий немыслимо продолжение жизни.
Образование высшее филологическое. В копирайтинге с 2012 г., также занимаюсь редактированием/размещением статей. Увлечения — психология и кулинария.
Источник
Типы питания прокариот
Прокариоты характеризуются многообразием типов питания в отличие от растений (автотрофы) и животных (ге- теротрофы). Для характеристики типов питания используют три критерия: источник углерода, источник энергии, донор электронов (водорода). По источнику углерода делят на ав- тотрофы и гетеротрофы, по источнику энергии – на фото- трофы (используют солнечный свет) и хемотрофы (получа- ют энергию за счет окислительно-восстановительных реак- ций), по донору – электроны (водорода) на литотрофы (неорганические соединения Н2, NH3, H2S, Fe2 + , CO и др.) и органотрофы (используют в качестве донора электрона ор- ганические соединения). Таким образом, выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы, хемолитоавтотрофы, хемоорганоге- теротрофы.
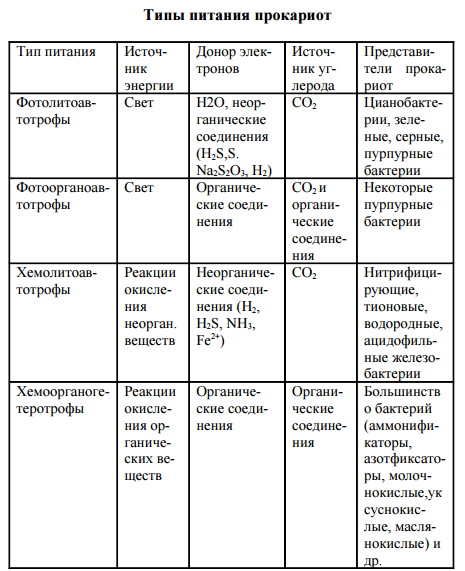
Фотолитоавтотрофы. Бактериальный фотосинтез (фоторедукция). В 1931 г. К. ван Ниль впервые доказал способность бактерий к фотосинтезу. В основе бактериального фотосинтеза лежит превращение световой энергии, поглощаемой фотосинтетическим пигментом, в биохимическую энергию макроэргических связей (АТФ) и далее использо- вание этой энергии для восстановления углекислого газа в процессе биосинтеза. У этих организмов есть пигмент бак- териохлорофилл. В клетках всех фотосинтезирующих бактерий содержатся фотосинтетические пигменты. К ним относятся особые хлорофиллы, получившие название бактериохлорофиллов а, в, с, d, и каротиноиды. По строению бактерио- хлорофиллы близки к хлорофиллу а растений. Так, бакте- риохлорофилл а отличается от хлорофилла а растений тем, что в первом его пирольном кольце в положении 2 стоит ацетильная группа СН3-СО- вместо обычной винильной группы СН2=СН-, а второе пирольное кольцо восстановле- но и содержит на два атома водорода больше. Содержание в клетке фотосинтезирующих прокариот четырех видоизменений бактериохлорофилла обусловли- вает более широкую полосу спектра поглощения световой энергии по сравнению с хлорофиллом растений. Предел спектра поглощения хлорофилла растений лежит в области 700-780 нм, в то время как фотосинтезирующие бактерии используют световую энергию длинноволновой части спектра до 1100 нм. Разница в спектрах поглощения хлоро- филлов растений и бактериохлорофиллов фотосинтезиру- ющих бактерий расширяет экологические ниши последних и позволяет фотосинтезирующим бактериям развиваться в водоемах под слоем водорослей. Помимо бактериохлорофиллов, в клетках фотосинтезирующих бактерий открыты более 20 дополнительных каро- тиноидных пигментов. Особенно разнообразны каротинои- ды в клетках пурпурных бактерий. Каротиноиды фотосин- тезирующих бактерий поглощают световую энергию ко- ротковолновой видимой области спектра с длиной волны 400-550 нм и передают эту энергию на бактериохлоро- филл. В клетке прокариот фотосинтетические пигменты нахо- дятся на инвагинациях ЦПМ – хроматофорах, визикулах (пузырьках), трубочках, тиллакоидах, ламеллах. Ван Ниль назвал этот процесс бактериальным фотосин- тезом, или фоторедукцией.
Для восстановления одной молекулы углекислого газа требуется 1 молекула АТФ (у растений 4 АТФ). В том и другом случае идет восстановление углекислого газа до углеводов. В Определителе Берджи (1974) фотосинтезирующие бактерии представлены тремя семействами (см. выше).
Фотоорганоавтотрофы Фотоорганоавтотрофы представлены немногочисленным семейством Rhodospirillaceae, включающим три рода: Rhodospirillum, Rhodopseudomonas и Rhodomicrobium. Бактерии-фотоорганоавтотрофы способны перестраивать свой обмен и одинаково успешно развиваться как на свету, так и в темноте, переходя соответственно от ана- эробного образа жизни к аэробному. На свету они ведут себя как фотоорганоавтотрофы: усваивая углекислый газ, они восстанавливают его в процессе фотосинтеза до углевода. В качестве доноров электронов несерные пурпурные бактерии используют различные органические вещества — сахара, спирты, органические кислоты, аминокислоты. Характер использования органических веществ различными фотосинтезирующими бактериями существенно раз- личается. Чаще всего органическое вещество выполняет единственную функцию донора электронов при фотоасси- миляции углекислого газа. Однако в некоторых случаях органическое вещество используется фотосинтезирующими бактериями не только в качестве донора электронов, но одновременно и как источник углерода. Попадая в темноту, пурпурные бактерии переходят к хемоорганогетеротрофному типу питания. При этом энергию для процессов жизнедеятельности они получают за счет реакций окисления органического субстрата по циклу Кребса. Непосредственным источником углерода и донором электронов для них являются органические соедине- ния субстрата. В клетках бактерий этой группы имеется универсальный набор дыхательных ферментов (НАД- и ФАД-дегидрогеназы и цитохромы), обеспечивающих им возможность перехода от анаэробного образа жизни на свету к аэробному в темноте и, соответственно, от авто трофного типа питания к гетеротрофному. Таким образом, среди разных групп прокариот нет, да и не может быть, резкой границы между различными типами питания.
ХемолитоавтотрофыХемолитоавтотрофы представлены микроорганизмами, способными в качестве основного источника углерода усваивать углекислый газ и синтезировать в клетке органические соединения, используя энергию реакций окисления неорганического субстрата. Для хемолитоавтотрофов неорганические вещества субстрата выступают донорами электронов в реакциях энергетического метаболизма и в процессе хемоассимиляции углекислого газа. Заслуга открытия процесса хемосинтеза принадлежит С. Н. Виноградскому. Он определил химизм как жизнь без органики. Им впервые была показана возможность образования органических веществ из неорганических в клетках бактерий, помимо процесса фотосинтеза. К хемолитоавтотрофам относится большинство видов нитрифицирующих, тионовых бактерий, некоторые виды из группы одноклеточных железобактерий и водородные бактерии. Хемолитоавтотрофные бактерии характеризуются специфичностью в отношении использования окисляемого субстрата. Нитрифицирующие бактерии для процесса хемоассимиляции углекислого газа получают энергию от окисления аммиака и нитритов. Процесс нитрификации проходит в две фазы. Первая фаза заключается в аэробном окислении аммиака до нитритов нитрозными бактериями родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira: NН3 + 11 /202 = HN02 + Н2О + 274,7 кДж. Вторая фаза нитрификации предусматривает окисление нитритов в нитраты нитратными бактериями родов Nitrobacter, Nitrospina, Nitrococcus: HN02 + 11 /202 =НNО3 + 87,5 кДж. При процессах окисления неорганического субстрата выделяется сравнительно небольшое количество энергии, и усваивается она клеткой с низким КПД, всего 5-10%. Поэтому для получения энергии на процессы жизнедеятельности нитрифицирующим бактериям приходится перерабатывать огромное количество субстрата. К хемолитоав- тотрофам относится большинство видов тионовых бактерий: Thiobacillus denitrificans, Т. thiooxidans, Т. acidophilus, Thiomicrospira pelophila и др. Они ведут процесс хемоасси- миляции углекислого газа, получая энергию за счет окисления восстановленных или частично восстановленных соединений серы, сероводорода, элементарной серы, тиосульфата и сульфита. Конечным продуктом окисления обычно является сульфат.
Некоторые виды тионовых бактерий (Thiobacillus ferrooxidans) способны получать энергию за счет окисления не только соединений серы, но и закисного железа (Fe2+) (разрушение труб). Хемолитоавтотрофный тип питания характерен для некоторых одноклеточных ацидофильных железобактерий – Leptospirillum ferrooxidans, Thiobacillus ferrooxidans и представителей рода Sulfolobus. Оптимальный рост ацидофильных железобактерий наблюдается при рН ниже 4,5 (2-3). Они используют энергию окисления Fe2+ до Fе3+ для ассимиляции С02, который служит основным или единственным источником углерода. Реакции окисления железа сопровождаются незначительным выделением энергии, поэтому железобактерии перерабатывают большие количества субстрата: 2Fe2+ + 1/202 + 2Н+ = 2FеЗ+ + Н2О + 33 кДж. Хемолитоавтотрофные бактерии в природе являются геологическими агентами. Они принимают участие в процессах образования полезных ископаемых и осуществляют важнейшие звенья круговорота азота, серы, железа. К факультативным хемолитоавтотрофам относятся водородные бактерии, являющиеся представителями 20 различных родов – Рsеudоmоnаs, Аlcаligеnеs, Nocardia и др. Они способны осуществлять ферментативное окисление водорода кислородом воздуха с образованием воды. Водородные бактерии ферментом дегидрогеназой активируют молекулярный водород и далее используют его для получения энергии и в качестве донора электронов для восстановления углекислого газа до углевода: 6Н2 + 202 + С02 = (СН2О) + 5Н2О. Являясь факультативными хемолитоавтотрофами, водородные бактерии в качестве источника энергии и углеро- да способны использовать и различные органические со- единения (сахара, органические кислоты, спирты). Таким образом, водородные бактерии могут служить еще одним 87 примером возможного перехода микробной клетки от одного типа питания к другому – от хемолитоавтотрофного к хемоорганогетеротрофному. Бактерии, переходящие от одного типа питания к другому, получили название миксо- трофов.
Хемоорганогетеротрофы. К хемоорганогетеротрофам относится большинство прокариот. Источником углерода для них являются самые разнообразные органические соединения. Энергию для жизнедеятельности они получают за счет окислительно- восстановительных реакций органического субстрата, и донором электронов в реакциях метаболизма также выступают различные органические вещества. Хемоорганогетеротрофы наиболее широко распространены в природе. Им принадлежит роль санитаров нашей планеты, так как они ведут процессы минерализации самых разнообразных, подчас сложных органических веществ. Помимо органических соединений как источника углерода, хемоорганогетеротрофы нуждаются в углекислом газе для реакций карбоксилирования промежуточного обмена. Хемоорганогетеротрофные микроорганизмы подразделяют на сапрофитов и паразитов. Сапрофиты потребляют органические вещества опада. Паразиты живут за счет органических веществ живой клетки. Выделяют факультативных и облигатных паразитов. Факультативные паразиты развиваются на обычных органических средах, но, попадая в клетку-хозяина, переходят к паразитическому образу жизни. К ним относится большинство патогенных бактерий, вызывающих заболевания человека, – возбудители пневмонии, менингита, гонореи, дизентерии, брюшного тифа, сибирской язвы, коклюша, туберкулеза и др. Облигатные (строгие) паразиты развиваются исключительно за 88 счет органических веществ клетки-хозяина. Типичным примером облигатных паразитов являются риккетсии и вирусы. В основе всех вышерассмотренных типов питания прокариот лежат различные сочетания их энергетического и конструктивного метаболизма. Всем типам питания соот- ветствуют определенные группы прокариотных организ- мов. Основной массе бактерий присущ один вполне определенный (облигатный) тип питания. Однако среди многообразного мира прокариот встречается много видов бактерий, способных переходить от одного типа питания к другому. Так, среди цианобактерий, зеленых и пурпурных бактерий имеются виды, переходящие от фотолитоавто- трофного типа питания к фотоорганогетеротрофному. Некоторые виды обширного рода Thiobacillus способны переходить от хемолитоавтотрофного типа питания к хемоорга- ногетеротрофному. Эта особенность питания прокариот обусловливает широкие возможности существования микроорганизмов в различных условиях среды и значительно расширяет их экологические ниши.
Источник



