
- полуконсервативная репликация
- Смотреть что такое «полуконсервативная репликация» в других словарях:
- Репликация происходит перед делением клетки
- Биология. 11 класс
- § 16. Клеточный цикл. Репликация ДНК
- Полуконсервативная репликация
- СОДЕРЖАНИЕ
- Открытие [ править ]
- Модели репликации [ править ]
- Разделение и рекомбинация двухцепочечной ДНК [ править ]
- Скорость и точность [ править ]
- Приложения [ править ]
полуконсервативная репликация
Смотреть что такое «полуконсервативная репликация» в других словарях:
полуконсервативная репликация — Метод репликации молекулы ДНК, при которой каждая материнская цепь молекулы ДНК достраивает себе новую дочернюю цепь [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN semiconservative replication … Справочник технического переводчика
Полуконсервативная репликация — * паўкансерватыўная рэплікацыя * semiconservative replication тип репликации ДНК, при котором молекула делится продольно, каждая половина сохраняется и служит матрицей для образующейся новой нити. Термин был введен, когда точный процесс… … Генетика. Энциклопедический словарь
Репликация полуконсервативная — * рэплікацыя паўкансерватыўная * semi conservative replication процесс репликации, при котором дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь синтезируется вновь. Такой же процесс повторяется при образовании… … Генетика. Энциклопедический словарь
репликация θ-типа — репликация θ типа θ репликация Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования «вздутия», видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Справочник технического переводчика
θ-репликация — θ type replication репликация θ типа, θ репликация. Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования вздутия , видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Молекулярная биология и генетика. Толковый словарь.
Семиконсервативная репликация — * семікансерватыўная рэплікацыя * semiconservative replication … Генетика. Энциклопедический словарь
генетический код — Долгое время наследственные механизмы были предметом внимания генетиков, но природа молекул, переносящих информацию от одного индивида к другому, оставалась неведомой. Была известна роль макромолекул протеина и нуклеиновых кислот в этом… … Западная философия от истоков до наших дней
История науки — История науки это исследование феномена науки в его истории. Наука, в частности, представляет собой совокупность эмпирических, теоретических и практических знаний о Мире, полученных научным сообществом. Поскольку с одной стороны наука… … Википедия
semiconservative replication — semiconservative replication. См. полуконсервативная репликация. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
5 -гидроксиметилцитозин 5 -гМц — 5 гидроксиметилцитозин, 5 гМц * 5 гідроксіметылцытазін * 5 hydroxymethyl cytosine or 5 hMC пиримидиновое основание, замещающее цитозин в ДНК четных колифагов. 5 ГМЦ комплементарен гуанину. Считается, что фагоспецифичная ДНКаза бактериофага… … Генетика. Энциклопедический словарь
Источник
Репликация происходит перед делением клетки
Репликация (синтез) ДНК происходит не беспорядочно, а в строго определенный период жизни клетки. Всего выделяют 4 фазы клеточного цикла: митоз (М), синтетическую (S), пресинтетическую (G1, от англ. gap – интервал), постсинтетическую (G2).
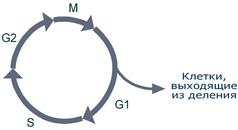
Фазы клеточного цикла
Важное участие в регуляции смены фаз клеточного цикла занимают циклины – белки массой 35-90 кДа, уровень которых меняется в ходе клеточного цикла. По функции циклины – это активаторные субъединицы ферментов циклин-зависимых киназ (ЦЗК). Активные комплексы циклин-ЦЗК фосфорилируют внутриклеточные белки, изменяя их активность. Этим обеспечивается продвижение по клеточному циклу.
Синтез (репликация, удвоение) ДНК происходит в S-фазу клеточного цикла, когда клетка готовится к делению. Механизм репликации, как установили Мэтью Мезельсон и Франклин Сталь в 1957 г, полуконсервативный, т.е. на каждой нити материнской ДНК синтезируется дочерняя копия.
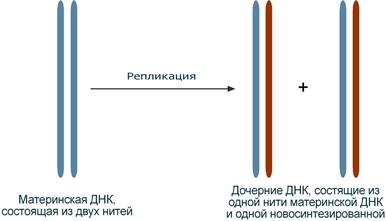
Схема полуконсервативного способа синтеза ДНК
Как любой матричный биосинтез, репликация требует наличия нескольких компонентов:
- матрица – в ее роли выступает материнская нить ДНК,
- растущая цепь – дочерняя нить ДНК,
- субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ,
- источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ,
- ферменты.
Синтез ДНК начинается в определенных участках, получивших название точка ori (англ. origin – начало). На каждой ДНК млекопитающих точек ori насчитывается до 100. Репликация распространяется от этих участков в обе стороны по нитям ДНК с образованием репликативных «пузырей«. В каждом таком «пузыре» имеются две репликативные «вилки», в которых происходит расплетание, раскручивание и непосредственный синтез ДНК. При этом репликативные вилки удаляются друг от друга. В целом репликация всех ДНК у эукариот заканчивается за 9 часов.
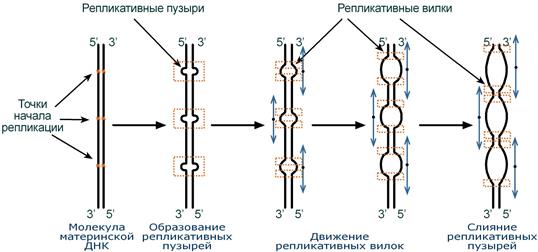
Схема возникновения репликативных пузырей и движения репликативных вилок
(нити дочерней ДНК не показаны). Стрелки — направление движения репликативных вилок.
Синтез новой цепи ДНК идет в направлении от 5′-конца к 3′-концу, т.е. 5′-конец новой ДНК остается свободным, каждый следующий нуклеотид своей 5′-гидроксильной группой присоединяется к 3′-гидроксильной группе предыдущего нуклеотида со скоростью порядка 100 штук в секунду.
В репликативной вилке в направлении 5’→3′ непрерывно (т.е. обычным заурядным присоединением последующих нуклеотидов к предыдущим через С3 и С5) синтезируется только одна нить, а именно та, для которой направление синтеза совпадает с направлением движения репликативной вилки и соответствует направлению материнской нити 3’→5′. По мере расплетания ДНК и движения репликативной вилки на этой материнской нити открываются участки, где возможно безостановочное удлинение ведущей дочерней нити.
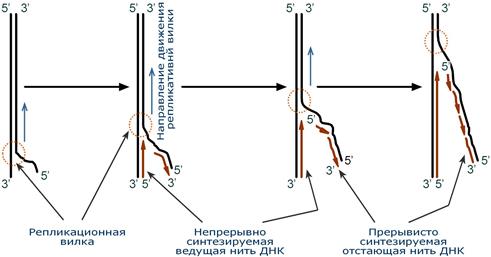
Схема движения репликативной вилки и синтеза ведущей и отстающей нитей ДНК
Направление 5’→3′ для другой дочерней нити ДНК противоположно движению репликативной вилки. Поэтому синтез этой отстающей нити (в направлении 5’→3′) возможен только после расплетания части ДНК и освобождения участка для синтеза.
Таким образом, синтез дочерней ДНК идет фрагментарно. По имени японского исследователя синтезируемые на отстающей цепи отрезки ДНК назвали фрагменты Оказаки.
Источник
Биология. 11 класс
§ 16. Клеточный цикл. Репликация ДНК
Клеточный цикл . Все новые клетки образуются путем деления уже существующих. Этот принцип, как уже отмечалось, сформулировал Р. Вирхов еще в середине XIX в. Деление клеток обеспечивает непрерывность существования жизни на нашей планете. Именно благодаря ему осуществляются различные способы бесполого и полового размножения организмов. В основе процессов роста, регенерации и индивидуального развития многоклеточных организмов также лежит деление клеток.
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
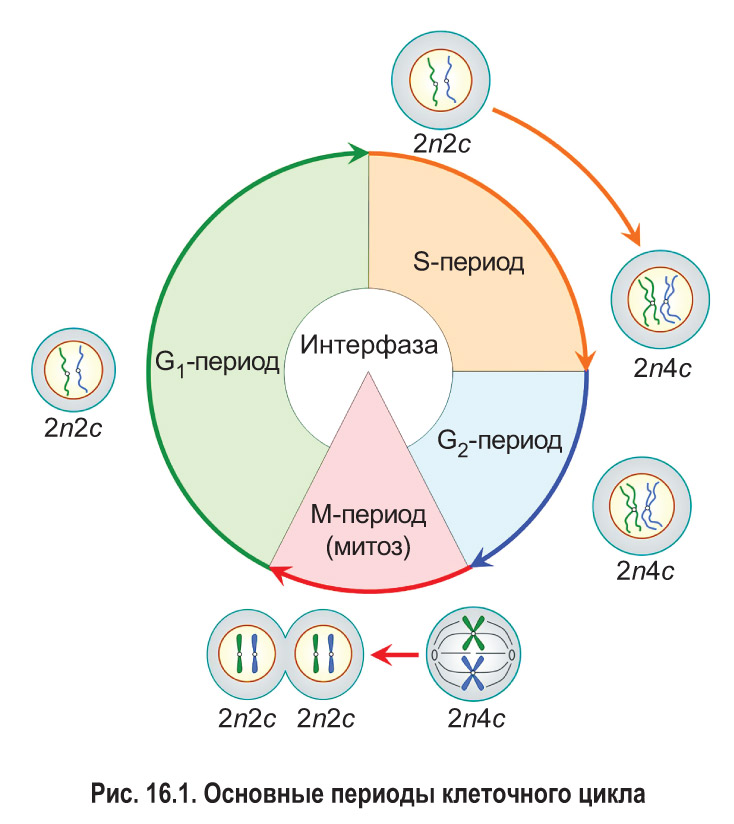 Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Итак, интерфаза обычно включает три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На протяжении всей интерфазы хромосомы не спирализованы. Они располагаются в ядре клетки в виде хроматина .
После завершения интерфазы начинается деление клетки. Основным способом деления клеток эукариот является митоз, поэтому данный этап клеточного цикла обозначают как М-период. При митозе происходит спирализация хроматина . Это приводит к формированию компактных двухроматидных хромосом. После этого сестринские хроматиды каждой хромосомы отделяются друг от друга и затем попадают в разные дочерние клетки. Значит, дочерние клетки, образовавшиеся в результате митоза и вступающие в новый клеточный цикл, имеют набор 2n2c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Источник
Полуконсервативная репликация
Полуконсервативная репликация описывает механизм репликации ДНК во всех известных клетках. Репликация ДНК происходит в нескольких точках репликации вдоль цепи матрицы ДНК. Поскольку двойная спираль ДНК разматывается геликазой , репликация происходит отдельно на каждой цепи матрицы в антипараллельных направлениях. Этот процесс известен как полуконсервативная репликация, потому что создаются две копии исходной молекулы ДНК. [1] Каждая копия содержит одну исходную цепочку и одну вновь синтезированную цепочку. Структура ДНК (расшифрованная Джеймсом Д. Уотсоном и Фрэнсисом Крикомв 1953 г.) предположил, что каждая цепь двойной спирали может служить шаблоном для синтеза новой цепи. Неизвестно, как вновь синтезированные цепи объединяются с цепями-матрицами с образованием двух двойных спиральных молекул ДНК. [2] [3]
СОДЕРЖАНИЕ
Открытие [ править ]
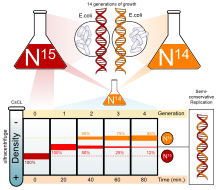
Было проведено множество экспериментов, чтобы определить, как реплицируется ДНК. Полуконсервативная модель была разгадана Н. Кольцова , а затем при поддержке эксперимента Мезельсон-Stahl ,. [3] [4] , которые подтвердили , что ДНК реплицируется пол-консервативна путем проведения эксперимента с использованием двух изотопов : азот-15 ( 15
N
) и азот-14 ( 14
N
). Когда 14
N
был добавлен к тяжелому 15
N
— 15
N
ДНК, гибрид 15
N
— 14
N
был замечен в первом поколении. После второго поколения гибрид остался, но легкая ДНК ( 14
N
— 14
N
) тоже было замечено. Это указывало на полуконсервативную репликацию ДНК. Этот режим репликации ДНК позволял каждой дочерней цепи оставаться связанной со своей цепочкой-матрицей. [5]
Модели репликации [ править ]
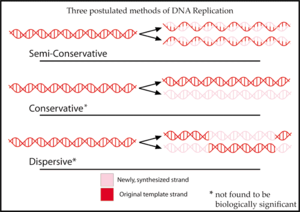
Полуконсервативная репликация получила свое название от того факта, что этот механизм транскрипции был одной из трех моделей, первоначально предложенных [2] [3] для репликации ДНК :
- Полуконсервативная репликация даст две копии, каждая из которых содержит одну из исходных цепей ДНК и одну новую цепочку. [2] Полуконсервативная репликация полезна для восстановления ДНК. Во время репликации новая цепь ДНК приспосабливается к модификациям, внесенным в цепочку-матрицу. [6]
- Консервативная репликация оставит две исходные цепи ДНК-матрицы вместе в двойной спирали и создаст копию, состоящую из двух новых цепей, содержащих все новые пары оснований ДНК. [2]
- Дисперсионная репликация создаст две копии ДНК, каждая из которых содержит отдельные участки ДНК, состоящие либо из обеих исходных цепей, либо из обеих новых цепей. [2] Изначально считалось, что цепи ДНК разрываются на каждой десятой паре оснований, чтобы добавить новую матрицу ДНК. В конце концов, вся новая ДНК будет образовывать двойную спираль после многих поколений репликации. [7]
Разделение и рекомбинация двухцепочечной ДНК [ править ]
Для полуконсервативной репликации двойную спираль ДНК необходимо разделить, чтобы новая цепь матрицы могла быть связана с комплементарными парами оснований. Топоизомераза — это фермент, который способствует распаковке и рекомбинации двойной спирали. В частности, топоизомераза предотвращает сверхспирализацию двойной спирали или ее слишком тугое наматывание. Три топоизомеразы ферменты участвуют в этом процессе: Тип IA топоизомеразы , тип IB топоизомеразы и Type II топоизомеразы . [8] Топоизомераза типа I раскручивает двухцепочечную ДНК, в то время как топоизомераза типа II разрывает водородные связи, связывающие комплементарные пары оснований ДНК. [7]
Скорость и точность [ править ]
Скорость полуконсервативной репликации ДНК в живой клетке сначала измеряли как скорость удлинения цепи ДНК фага Т4 в инфицированной фагом E. coli . [9] В период экспоненциального увеличения ДНК при 37 ° C скорость удлинения цепи составляла 749 нуклеотидов в секунду. Частота мутаций на пару оснований за цикл репликации во время синтеза ДНК фага Т4 составляет 2,4 × 10 −8 . [10] Таким образом, полуконсервативная репликация ДНК происходит быстро и точно.
Приложения [ править ]
Полуконсервативная репликация дает ДНК множество преимуществ. Это быстро, точно и позволяет легко восстанавливать ДНК. Он также отвечает за фенотипическое разнообразие у нескольких видов прокариот. [11] Процесс создания вновь синтезированной цепи из цепи-шаблона позволяет метилировать старую цепь в отдельное время от новой цепи. Это позволяет репаративным ферментам проверять новую цепь и исправлять любые мутации или ошибки. [6]
ДНК может иметь способность активировать или деактивировать определенные области на вновь синтезированной цепи, что позволяет изменять фенотип клетки. Это может быть выгодно для клетки, потому что ДНК может активировать более благоприятный фенотип, чтобы помочь в выживании. Благодаря естественному отбору более благоприятный фенотип будет сохраняться у всего вида. Это порождает идею наследования или того, почему одни фенотипы наследуются над другими. [6]
Источник



