
Полуконсервативный способ репликации




![]()
![]()
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.
Гипотеза Уотсона-Крика была проверена с помощью опытов, выполненных М.Мезельсоном и Ф.Сталем в 1957 г. Клетки E.coli выращивали в течение ряда поколений в среде, содержащей в качестве источника азота хлористый аммоний NH4Cl, в котором обычный изотоп [ 14 N] был заменен на «тяжелый» изотоп [ 15 N]. Вследствие этого все соединения клеток, имеющие в своем составе азот, в том числе и азотистые основания ДНК, оказались обогащенными изотопом [ 15 N]. Плотность ДНК, выделенной из этих клеток, была выше плотности нормальной [ 14 N] ДНК. Смесь «тяжелой» [ 15 N] и «легкой» [ 14 N] ДНК удалось разделить методом центрифугирования в концентрированном растворе хлористого цезия. Поскольку [ 15 N] ДНК чуть тяжелее, чем [ 14 N] ДНК, полоса, в которой она достигает равновесия в градиенте СsCl, расположена ближе ко дну пробирки, чем полоса с [ 14 N] ДНК (рис.7)
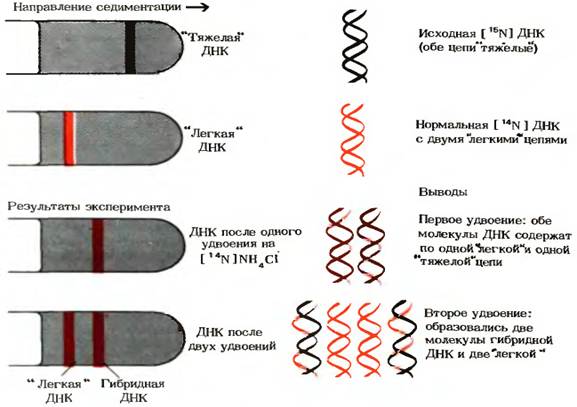
Рис. 7 Результаты эксперимента Мезельсона-Сталя
Мезельсон и Сталь перенесли клетки E.coli, росшие на среде с изотопом [ 15 N] и содержащие «тяжелые» цепи ДНК, на свежую среду с обычным изотопом [ 14 N]. На этой среде клетки E.coli выращивали в течение времени, необходимого для удвоения клеток. Затем из этих клеток выделяли ДНК и анализировали ее плотность с помощью описанного выше метода седиментации (осаждения ДНК в растворе СsCl). В градиенте СsCl была обнаружена лишь одна полоса ДНК, плотность которой оказалась средней между плотностью нормальной «легкой» [ 14 N] ДНК и плотностью «тяжелой» [ 15 N] ДНК (рис. ). Это подтвердило, что двухцепочечная ДНК дочерних клеток содержит одну старую 15 N – цепь от родительской ДНК и одну новую 14 N – цепь.
Если выделить ДНК из клеток, которые прошли два цикла удвоения на среде с [ 14 N], то она разделится на две полосы: одна с плотностью, соответствующей плотности нормальной «легкой» ДНК, а другая с плотностью «гибридной» ДНК, наблюдавшейся после первого удвоения клеток. На основании этих данных Мезельсон и Сталь пришли к выводу, что в строгом соответствии с гипотезой Уотсона-Крика каждый дочерний дуплекс ДНК после двух циклов удвоения клеток содержал одну родительскую и одну новообразованную цепь ДНК. Такой механизм репликации назвали полуконсервативным. Полученные результаты полностью исключили консервативный способ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсон и Сталя позволил также отвергнуть дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из коротких участков как родительской, так и новообразованнй ДНК, соединенных между собой случайным образом.
Источник
Биология. 11 класс
§ 16. Клеточный цикл. Репликация ДНК
Клеточный цикл . Все новые клетки образуются путем деления уже существующих. Этот принцип, как уже отмечалось, сформулировал Р. Вирхов еще в середине XIX в. Деление клеток обеспечивает непрерывность существования жизни на нашей планете. Именно благодаря ему осуществляются различные способы бесполого и полового размножения организмов. В основе процессов роста, регенерации и индивидуального развития многоклеточных организмов также лежит деление клеток.
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
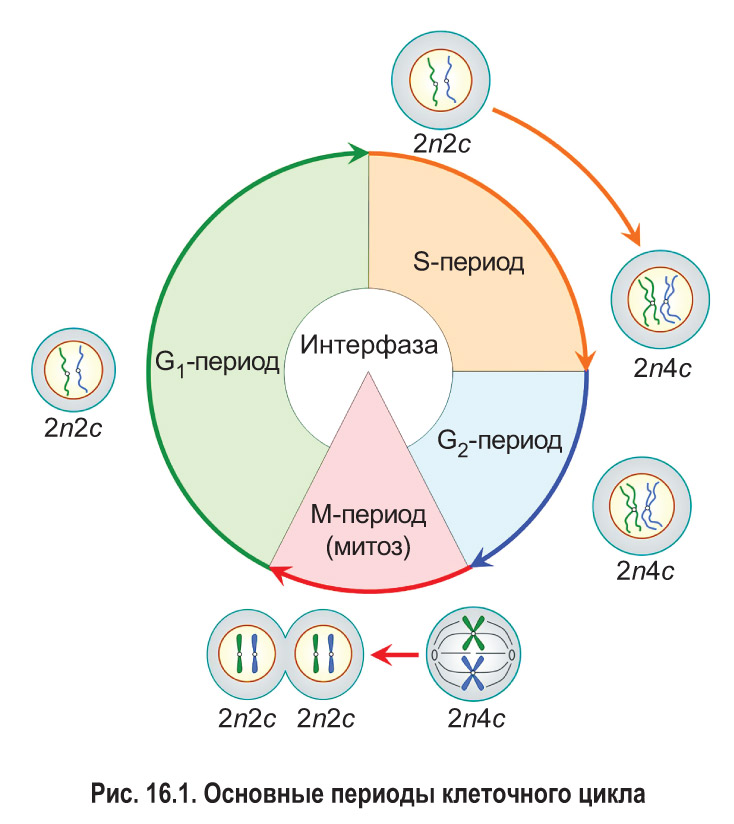 Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Итак, интерфаза обычно включает три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На протяжении всей интерфазы хромосомы не спирализованы. Они располагаются в ядре клетки в виде хроматина .
После завершения интерфазы начинается деление клетки. Основным способом деления клеток эукариот является митоз, поэтому данный этап клеточного цикла обозначают как М-период. При митозе происходит спирализация хроматина . Это приводит к формированию компактных двухроматидных хромосом. После этого сестринские хроматиды каждой хромосомы отделяются друг от друга и затем попадают в разные дочерние клетки. Значит, дочерние клетки, образовавшиеся в результате митоза и вступающие в новый клеточный цикл, имеют набор 2n2c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Источник
Полуконсервативный способ репликации днк схема

Biology exam запись закреплена
Экзаменационный вопрос №5.Способы репликации ДНК: консервативный, полуконсервативный, дисперсионный. Опыты Мезельсон и Сталь
В 1957 г. Метью Мезелсон и Франклин Сталь поставили один из самых красивых биологических экспериментов, исследуя механизмы репликации ДНК. Для определения способа репликации ДНК необходимо четко различать материнские и дочерние молекулы. Мезелсон и Сталь выращивали бактерии кишечной палочки (Escherichia coli) на среде, содержащей в качестве источника азота его тяжелый изотоп – 15N. Молекула ДНК содержит большое количество атомов азота, при делении ДНК азот для новых цепочек берется из внешней среды. Тяжелый изотоп азота включался в состав молекулы ДНК и служил надежной меткой. Для того чтобы пометить практически всю бактериальную ДНК, необходимо было культивировать E. coli на такой среде в течение как минимум 12 поколений.
Мезелсон и Сталь длительное время культивировали E. coli на среде с 15N. После этого бактерии быстро переносили на среду, содержащую более легкий изотоп азота 14N. Благодаря тому, что клетки E. coli культивировались на двух различных средах, в состав молекул их ДНК входили оба изотопа азота: 14N и 15N. Отличить такие молекулы друг от друга можно было по плотности, поскольку масса нуклеотидов молекулы ДНК, содержащих 15N больше, по сравнению с обычной молекулой. ДНК бактериальных клеток, выращенных на среде с 15N, имела плотность 1,724 г/см3, а ДНК клеток, выращенных на среде с 14N – 1,710 г/см3.
После переноса культуры E. coli с одной среды на другую через каждое поколение Мезелсон и Сталь отбирали пробы. Контролем служила бактериальная культура, содержавшаяся на среде с 15N. Из каждой пробы бактериальных клеток путем центрифугирования извлекали ДНК. После этого ее смешивали с раствором хлористого цезия (CsCl) плотностью 1,7 г/см3, и вновь центрифугировали с очень высокой скоростью в течение нескольких дней. В результате осаждения молекул CsCl его раствор приобретал градиент плотности от 1,65 г/см3 в верхней части пробирки до 1,8 г/см3 у ее дна. В соответствии с этим, молекулы ДНК концентрировались в строго определенной области: у дна, в центре или в верхней части пробирки. Локализация ДНК устанавливалась на спектрофотометре, поскольку было известно, что она поглощает лучи с длиной волны 260 нм. Этот метод Мезелсон и Сталь разработали и опубликовали совместно с Джермом Виноградом (1913–1976) в 1957 г., непосредственно перед проведением эксперимента по репликации ДНК. пределяя плотность ДНК в каждой из проб, Мезелсон и Сталь обнаружили, что спустя одно поколение после переноса культуры E. coli со среды с 15N на среду с 14N плотность ДНК была промежуточной между 15N-ДНК и 14N-ДНК. Спустя два поколения половина бактериальных клеток содержала ДНК с легким изотопом азота (14N), а другая половина – такую же, как и в предыдущем поколении, ДНК промежуточной плотности. Через три поколения на среде с 14N в ¾ клеток содержалась легкая ДНК, а ¼ часть клеток сохраняла ДНК промежуточной плотности. Т. о., соотношение между числом генераций и распределением плотности ДНК точно соответствовало полуконсервативному типу репликации.
месте с тем, из гипотезы полуконсервативной репликации следовало, что ДНК с промежуточной плотностью между 15N-ДНК и 14N-ДНК должна быть гибридной. Это значит, что одна из ее цепей должна содержать только тяжелый изотоп азота (15N), а другая – только легкий (14N). Проверяя это предположение, Мезелсон и Сталь нагревали полученную ими ДНК промежуточной плотности в течение 30 минут при температуре 100 °С на водяной бане. При таких условиях двойная спираль молекулы ДНК денатурирует, образуя две отдельных цепочки, однако ковалентные связи между нуклеотидами в каждой из них не разрушаются. Проведя центрифугирование денатурировавшей ДНК в градиенте плотности хлористого цезия, Мезелсон и Сталь обнаружили, что в результате образовалось две фракции различной плотности. Плотность одной из них совпадала с плотностью молекул ДНК, содержащих тяжелый изотоп азота (15N), тогда как плотность другой была идентична плотности молекул ДНК с легким изотопом азота (14N). Из этого следовало, что молекула ДНК промежуточной плотности, образовавшаяся в первом поколении после переноса со среды с тяжелым изотопом азота на среду с легким изотопом, представляет собой гибридную молекулу. В ее состав входят две цепи – материнская, содержащая исключительно 15N и вновь синтезированная дочерняя, содержащая только 14N. Эти данные также подтвердили верность модели Уотсона–Крика и полуконсервативный характер репликации ДНК.
— Консервативная репликация:
Молекула ДНК служит матрицей для образования совершенно новой молекулы ДНК. В результате одна из образующихся клеток получает исходную молекулу ДНК, а другая – вновь синтезированную.
— Полуконсервативная репликация:
Две цепи исходной молекулы ДНК расходятся вследствие разрыва слабых водородных связей между азотистыми основаниями. Каждая из них служит матрицей для образования новой цепи, а возникающие между азотистыми основаниями водородные связи соединяют старую и новую цепи, восстанавливая целостность молекулы. В результате каждая новая клетка получает гибридную молекулу ДНК, состоящую из одной старой и одной новой цепи.
— Дисперсная репликация:
ДНК распадается на короткие фрагменты, используемые в качестве матриц для построения фрагментов двух новых молекул ДНК, которые затем каким-то образом соединяются между собой.
Источник



