Полуконсервативный способ репликации днк доказательства
Расшифровка строения ДНК – результат многолетнего труда различных ученых – физиков, химиков и биологов. Начало было положено медиком и химиком русского происхождения Ф. Левиным. В двадцатых годах XX века он установил, что нуклеиновые кислоты – полимеры, образованные из мономеров нуклеотидов, в состав которых входят пуриновые основания – аденин и гуанин, и пиримидиновые – тимин, цитозин и урацил. В эксперименте, проведенном совместно с русским физиологом Е. С. Лондоном, Левину удалось выделить ДНК. Через двадцать лет американский биохимик Эрвин Чаргафф показал, что количество оснований А и Т соответствует количеству Ц и Г. В 1953 г. американский химик Лайнус Полинг опубликовал статью, в которой теоретически доказывал, что минимум энергии в глобулярных белках достигается при конфигурации молекулы, свернутой в спираль. Ошибка Полинга заключалась в том, что он предположил спираль трехцепочной. Подтверждение спиральной модели строения ДНК было получено рентгенологами Морисом Уилкинсом и Розалиндой Франклин.
Для расшифровки результатов рассеяния рентгеновских лучей понадобилось разработать новые методы анализа, т. к. впервые изучалось рассеяние на спиралевидных структурах. Результаты экспериментов подтвердили предположения Полинга и позволили рассчитать шаг спирали. На основании этих работ в 1953 г. Джеймс Уотсон и Френсис Крик предложили модель двойной спирали молекулы ДНК.
К этому времени было доказано, что важнейшими функциями ДНК является хранение и передача наследственной информации от родителей к потомкам. Передача наследственной информации происходит в результате:
репликации родительской ДНК,
транскрипции генетической информации на матрицу РНК,
трансляции информации с РНК в белковую форму.
Было известно, что цепочки ДНК состоят из четырех оснований, обозначаемых А, Т, Ц, Г. Причем шариковые модели демонстрировали, что по чисто геометрическим причинам на противоположных цепочках друг напротив друга могут располагаться только пары Уотсон и Крик считали, что две цепочки ДНК достраиваются автоматически, т. к. одной цепочки достаточно для построения другой. Стоял вопрос о том, как происходит удвоение ДНК: делится ли исходная молекула на фрагменты, если делится, то на какие, и как из этих фрагментов получается новая молекула. Рассматривались три гипотезы.
Консервативная репликация. Молекула ДНК служит матрицей для образования совершенно новой молекулы ДНК. В результате одна из образующихся клеток получает исходную молекулу ДНК, а другая – вновь синтезированную.
Полуконсервативная репликация. Две цепи исходной молекулы ДНК расходятся вследствие разрыва слабых водородных связей между азотистыми основаниями. Каждая из них служит матрицей для образования новой цепи, а возникающие между азотистыми основаниями водородные связи соединяют старую и новую цепи, восстанавливая целостность молекулы. В результате каждая новая клетка получает гибридную молекулу ДНК, состоящую из одной старой и одной новой цепи.
Дисперсная репликация. ДНК распадается на короткие фрагменты, используемые в качестве матриц для построения фрагментов двух новых молекул ДНК, которые затем образом соединяются между собой.
В 1957 г. Метью Мезелсон и Франклин Сталь поставили один из самых красивых биологических экспериментов, исследуя механизмы репликации ДНК. Для определения способа репликации ДНК необходимо четко различать материнские и дочерние молекулы. Мезелсон и Сталь выращивали бактерии кишечной палочки () на среде, содержащей в качестве источника азота его тяжелый изотоп – 15 N. Молекула ДНК содержит большое количество атомов азота, при делении ДНК азот для новых цепочек берется из внешней среды. Тяжелый изотоп азота включался в состав молекулы ДНК и служил надежной меткой. Для того чтобы пометить практически всю бактериальную ДНК, необходимо было культивировать на такой среде в течение как минимум 12 поколений.
Мезелсон и Сталь длительное время культивировали на среде с 15 N. После этого бактерии быстро переносили на среду, содержащую более легкий изотоп азота 14 N. Благодаря тому, что клетки культивировались на двух различных средах, в состав молекул их ДНК входили оба изотопа азота: 14 N и 15 N. Отличить такие молекулы друг от друга можно было по плотности, поскольку масса нуклеотидов молекулы ДНК, содержащих 15 N больше, по сравнению с обычной молекулой. ДНК бактериальных клеток, выращенных на среде с 15 N, имела плотность 1,724 г/см 3 , а ДНК клеток, выращенных на среде с 14 N – 1,710 г/см 3 .
После переноса культуры с одной среды на другую через каждое поколение Мезелсон и Сталь отбирали пробы. Контролем служила бактериальная культура, содержавшаяся на среде с 15 N. Из каждой пробы бактериальных клеток путем центрифугирования извлекали ДНК. После этого ее смешивали с раствором хлористого цезия (CsCl) плотностью 1,7 г/см 3 , и вновь центрифугировали с очень высокой скоростью в течение нескольких дней. В результате осаждения молекул CsCl его раствор приобретал градиент плотности от 1,65 г/см 3 в верхней части пробирки до 1,8 г/см 3 у ее дна. В соответствии с этим, молекулы ДНК концентрировались в строго определенной области: у дна, в центре или в верхней части пробирки. Локализация ДНК устанавливалась на спектрофотометре, поскольку было известно, что она поглощает лучи с длиной волны 260 нм. Этот метод Мезелсон и Сталь разработали и опубликовали совместно с Джермом Виноградом в 1957 г., непосредственно перед проведением эксперимента по репликации ДНК.
Определяя плотность ДНК в каждой из проб, Мезелсон и Сталь обнаружили, что спустя одно поколение после переноса культуры со среды с 15 N на среду с 14 N плотность ДНК была промежуточной между и Спустя два поколения половина бактериальных клеток содержала ДНК с легким изотопом азота ( 14 N), а другая половина – такую же, как и в предыдущем поколении, ДНК промежуточной плотности. Через три поколения на среде с 14 N в ¾ клеток содержалась легкая ДНК, а ¼ часть клеток сохраняла ДНК промежуточной плотности. Т. о., соотношение между числом генераций и распределением плотности ДНК точно соответствовало полуконсервативному типу репликации.
Вместе с тем, из гипотезы полуконсервативной репликации следовало, что ДНК с промежуточной плотностью между и должна быть гибридной. Это значит, что одна из ее цепей должна содержать только тяжелый изотоп азота ( 15 N), а другая – только легкий ( 14 N). Проверяя это предположение, Мезелсон и Сталь нагревали полученную ими ДНК промежуточной плотности в течение 30 минут при температуре 100 °С на водяной бане. При таких условиях двойная спираль молекулы ДНК денатурирует, образуя две отдельных цепочки, однако ковалентные связи между нуклеотидами в каждой из них не разрушаются. Проведя центрифугирование денатурировавшей ДНК в градиенте плотности хлористого цезия, Мезелсон и Сталь обнаружили, что в результате образовалось две фракции различной плотности. Плотность одной из них совпадала с плотностью молекул ДНК, содержащих тяжелый изотоп азота ( 15 N), тогда как плотность другой была идентична плотности молекул ДНК с легким изотопом азота ( 14 N). Из этого следовало, что молекула ДНК промежуточной плотности, образовавшаяся в первом поколении после переноса со среды с тяжелым изотопом азота на среду с легким изотопом, представляет собой гибридную молекулу. В ее состав входят две цепи – материнская, содержащая исключительно 15 N и вновь синтезированная дочерняя, содержащая только 14 N. Эти данные также подтвердили верность модели и полуконсервативный характер репликации ДНК.
Результаты своего эксперимента Мезелсон и Сталь опубликовали годом позже. В дальнейшем, было проделано множество экспериментов по репликации ДНК различных организмов – от прокариот до эукариот. Во всех случаях репликация ДНК шла исключительно по полуконсервативному типу. Это позволило подтвердить справедливость модели молекулы ДНК на которой базируется вся современная генетика.
Источник
Полуконсервативный способ репликации
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.
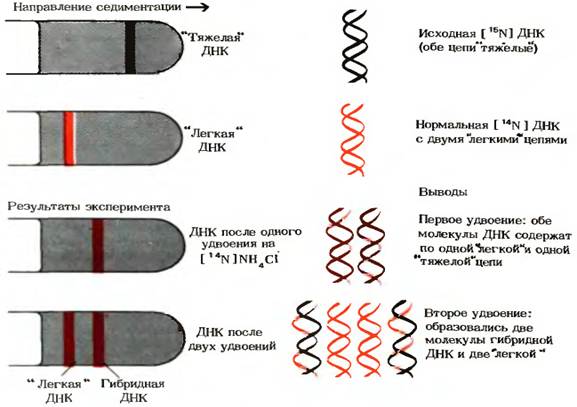
Гипотеза Уотсона-Крика была проверена с помощью опытов, выполненных М.Мезельсоном и Ф.Сталем в 1957 г. Клетки E.coli выращивали в течение ряда поколений в среде, содержащей в качестве источника азота хлористый аммоний NH4Cl, в котором обычный изотоп [ 14 N] был заменен на «тяжелый» изотоп [ 15 N]. Вследствие этого все соединения клеток, имеющие в своем составе азот, в том числе и азотистые основания ДНК, оказались обогащенными изотопом [ 15 N]. Плотность ДНК, выделенной из этих клеток, была выше плотности нормальной [ 14 N] ДНК. Смесь «тяжелой» [ 15 N] и «легкой» [ 14 N] ДНК удалось разделить методом центрифугирования в концентрированном растворе хлористого цезия. Поскольку [ 15 N] ДНК чуть тяжелее, чем [ 14 N] ДНК, полоса, в которой она достигает равновесия в градиенте СsCl, расположена ближе ко дну пробирки, чем полоса с [ 14 N] ДНК (рис.7)
Рис. 7 Результаты эксперимента Мезельсона-Сталя
Мезельсон и Сталь перенесли клетки E.coli, росшие на среде с изотопом [ 15 N] и содержащие «тяжелые» цепи ДНК, на свежую среду с обычным изотопом [ 14 N]. На этой среде клетки E.coli выращивали в течение времени, необходимого для удвоения клеток. Затем из этих клеток выделяли ДНК и анализировали ее плотность с помощью описанного выше метода седиментации (осаждения ДНК в растворе СsCl). В градиенте СsCl была обнаружена лишь одна полоса ДНК, плотность которой оказалась средней между плотностью нормальной «легкой» [ 14 N] ДНК и плотностью «тяжелой» [ 15 N] ДНК (рис. ). Это подтвердило, что двухцепочечная ДНК дочерних клеток содержит одну старую 15 N – цепь от родительской ДНК и одну новую 14 N – цепь.
Если выделить ДНК из клеток, которые прошли два цикла удвоения на среде с [ 14 N], то она разделится на две полосы: одна с плотностью, соответствующей плотности нормальной «легкой» ДНК, а другая с плотностью «гибридной» ДНК, наблюдавшейся после первого удвоения клеток. На основании этих данных Мезельсон и Сталь пришли к выводу, что в строгом соответствии с гипотезой Уотсона-Крика каждый дочерний дуплекс ДНК после двух циклов удвоения клеток содержал одну родительскую и одну новообразованную цепь ДНК. Такой механизм репликации назвали полуконсервативным. Полученные результаты полностью исключили консервативныйспособ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсон и Сталя позволил также отвергнуть дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из коротких участков как родительской, так и новообразованнй ДНК, соединенных между собой случайным образом.
Источник
полуконсервативная репликация
Смотреть что такое «полуконсервативная репликация» в других словарях:
полуконсервативная репликация — Метод репликации молекулы ДНК, при которой каждая материнская цепь молекулы ДНК достраивает себе новую дочернюю цепь [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN semiconservative replication … Справочник технического переводчика
Полуконсервативная репликация — * паўкансерватыўная рэплікацыя * semiconservative replication тип репликации ДНК, при котором молекула делится продольно, каждая половина сохраняется и служит матрицей для образующейся новой нити. Термин был введен, когда точный процесс… … Генетика. Энциклопедический словарь
Репликация полуконсервативная — * рэплікацыя паўкансерватыўная * semi conservative replication процесс репликации, при котором дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь синтезируется вновь. Такой же процесс повторяется при образовании… … Генетика. Энциклопедический словарь
репликация θ-типа — репликация θ типа θ репликация Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования «вздутия», видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Справочник технического переводчика
θ-репликация — θ type replication репликация θ типа, θ репликация. Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования вздутия , видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Молекулярная биология и генетика. Толковый словарь.
генетический код — Долгое время наследственные механизмы были предметом внимания генетиков, но природа молекул, переносящих информацию от одного индивида к другому, оставалась неведомой. Была известна роль макромолекул протеина и нуклеиновых кислот в этом… … Западная философия от истоков до наших дней
История науки — История науки это исследование феномена науки в его истории. Наука, в частности, представляет собой совокупность эмпирических, теоретических и практических знаний о Мире, полученных научным сообществом. Поскольку с одной стороны наука… … Википедия
semiconservative replication — semiconservative replication. См. полуконсервативная репликация. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
5 -гидроксиметилцитозин 5 -гМц — 5 гидроксиметилцитозин, 5 гМц * 5 гідроксіметылцытазін * 5 hydroxymethyl cytosine or 5 hMC пиримидиновое основание, замещающее цитозин в ДНК четных колифагов. 5 ГМЦ комплементарен гуанину. Считается, что фагоспецифичная ДНКаза бактериофага… … Генетика. Энциклопедический словарь