
- Полуконсервативный способ репликации
- Полуконсервативная репликация — Semiconservative replication
- СОДЕРЖАНИЕ
- Открытие
- Модели репликации
- Разделение и рекомбинация двухцепочечной ДНК
- Скорость и точность
- Приложения
- Биология. 11 класс
- § 16. Клеточный цикл. Репликация ДНК
- § 16. Клеточный цикл. Репликация ДНК
- Оглавление
Полуконсервативный способ репликации




![]()
![]()
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.
Гипотеза Уотсона-Крика была проверена с помощью опытов, выполненных М.Мезельсоном и Ф.Сталем в 1957 г. Клетки E.coli выращивали в течение ряда поколений в среде, содержащей в качестве источника азота хлористый аммоний NH4Cl, в котором обычный изотоп [ 14 N] был заменен на «тяжелый» изотоп [ 15 N]. Вследствие этого все соединения клеток, имеющие в своем составе азот, в том числе и азотистые основания ДНК, оказались обогащенными изотопом [ 15 N]. Плотность ДНК, выделенной из этих клеток, была выше плотности нормальной [ 14 N] ДНК. Смесь «тяжелой» [ 15 N] и «легкой» [ 14 N] ДНК удалось разделить методом центрифугирования в концентрированном растворе хлористого цезия. Поскольку [ 15 N] ДНК чуть тяжелее, чем [ 14 N] ДНК, полоса, в которой она достигает равновесия в градиенте СsCl, расположена ближе ко дну пробирки, чем полоса с [ 14 N] ДНК (рис.7)
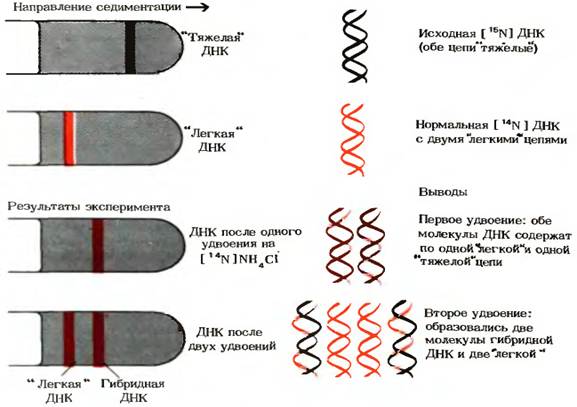
Рис. 7 Результаты эксперимента Мезельсона-Сталя
Мезельсон и Сталь перенесли клетки E.coli, росшие на среде с изотопом [ 15 N] и содержащие «тяжелые» цепи ДНК, на свежую среду с обычным изотопом [ 14 N]. На этой среде клетки E.coli выращивали в течение времени, необходимого для удвоения клеток. Затем из этих клеток выделяли ДНК и анализировали ее плотность с помощью описанного выше метода седиментации (осаждения ДНК в растворе СsCl). В градиенте СsCl была обнаружена лишь одна полоса ДНК, плотность которой оказалась средней между плотностью нормальной «легкой» [ 14 N] ДНК и плотностью «тяжелой» [ 15 N] ДНК (рис. ). Это подтвердило, что двухцепочечная ДНК дочерних клеток содержит одну старую 15 N – цепь от родительской ДНК и одну новую 14 N – цепь.
Если выделить ДНК из клеток, которые прошли два цикла удвоения на среде с [ 14 N], то она разделится на две полосы: одна с плотностью, соответствующей плотности нормальной «легкой» ДНК, а другая с плотностью «гибридной» ДНК, наблюдавшейся после первого удвоения клеток. На основании этих данных Мезельсон и Сталь пришли к выводу, что в строгом соответствии с гипотезой Уотсона-Крика каждый дочерний дуплекс ДНК после двух циклов удвоения клеток содержал одну родительскую и одну новообразованную цепь ДНК. Такой механизм репликации назвали полуконсервативным. Полученные результаты полностью исключили консервативный способ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсон и Сталя позволил также отвергнуть дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из коротких участков как родительской, так и новообразованнй ДНК, соединенных между собой случайным образом.
Источник
Полуконсервативная репликация — Semiconservative replication
Полуконсервативная репликация описывает механизм репликации ДНК во всех известных клетках. Репликация ДНК происходит в нескольких точках репликации вдоль цепи матрицы ДНК. Поскольку двойная спираль ДНК разматывается геликазой , репликация происходит отдельно на каждой цепи матрицы в антипараллельных направлениях. Этот процесс известен как полуконсервативная репликация, потому что создаются две копии исходной молекулы ДНК. Каждая копия содержит одну исходную цепочку и одну вновь синтезированную цепочку. Структура ДНК (расшифрованная Джеймсом Д. Уотсоном и Фрэнсисом Криком в 1953 г.) предполагала, что каждая цепь двойной спирали будет служить шаблоном для синтеза новой цепи. Неизвестно, как вновь синтезированные цепи объединяются с цепями-матрицами с образованием двух двойных спиральных молекул ДНК.
СОДЕРЖАНИЕ
Открытие
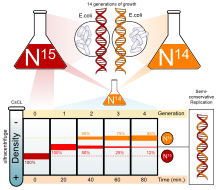
Было проведено множество экспериментов, чтобы определить, как реплицируется ДНК. Полуконсервативная модель была предвосхищена Николаем Кольцовым и позже подтверждена экспериментом Мезельсона-Шталя ., Который подтвердил, что ДНК реплицируется полуконсервативно путем проведения эксперимента с использованием двух изотопов : азота-15 ( 15
N
) и азот-14 ( 14
N
). Когда 14
N
был добавлен к тяжелому 15
N
— 15
N
ДНК, гибрид 15
N
— 14
N
был замечен в первом поколении. После второго поколения гибрид остался, но легкая ДНК ( 14
N
— 14
N
) тоже было замечено. Это указывало на полуконсервативную репликацию ДНК. Этот режим репликации ДНК позволял каждой дочерней цепи оставаться связанной со своей цепочкой-матрицей.
Модели репликации
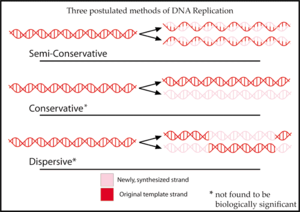
Полуконсервативная репликация получила свое название от того факта, что этот механизм транскрипции был одной из трех моделей, первоначально предложенных для репликации ДНК :
- Полуконсервативная репликация даст две копии, каждая из которых содержит одну из исходных цепей ДНК и одну новую цепочку. Полуконсервативная репликация полезна для восстановления ДНК. Во время репликации новая цепь ДНК приспосабливается к модификациям, внесенным в цепочку-матрицу.
- Консервативная репликация оставит две исходные цепи ДНК-матрицы вместе в двойной спирали и создаст копию, состоящую из двух новых цепей, содержащих все новые пары оснований ДНК.
- Дисперсионная репликация создаст две копии ДНК, каждая из которых содержит отдельные участки ДНК, состоящие либо из обеих исходных цепей, либо из обеих новых цепей. Первоначально считалось, что цепи ДНК разрываются на каждой десятой паре оснований, чтобы добавить новую матрицу ДНК. В конце концов, вся новая ДНК будет образовывать двойную спираль после многих поколений репликации.
Разделение и рекомбинация двухцепочечной ДНК
Для полуконсервативной репликации двойную спираль ДНК необходимо разделить, чтобы новая цепь матрицы могла быть связана с комплементарными парами оснований. Топоизомераза — это фермент, который способствует распаковке и рекомбинации двойной спирали. В частности, топоизомераза предотвращает сверхспирализацию двойной спирали или ее слишком тугое наматывание. Три топоизомеразы ферменты участвуют в этом процессе: Тип IA топоизомеразы , тип IB топоизомеразы и Type II топоизомеразы . Топоизомераза типа I раскручивает двухцепочечную ДНК, в то время как топоизомераза типа II разрывает водородные связи, связывающие комплементарные пары оснований ДНК.
Скорость и точность
Скорость полуконсервативной репликации ДНК в живой клетке сначала измеряли как скорость удлинения цепи ДНК фага Т4 в инфицированной фагом E. coli . В период экспоненциального увеличения ДНК при 37 ° C скорость удлинения цепи составляла 749 нуклеотидов в секунду. Частота мутаций на пару оснований за раунд репликации во время синтеза ДНК фага Т4 составляет 2,4 × 10 −8 . Таким образом, полуконсервативная репликация ДНК происходит быстро и точно.
Приложения
Полуконсервативная репликация дает ДНК множество преимуществ. Это быстро, точно и позволяет легко восстанавливать ДНК. Он также отвечает за фенотипическое разнообразие у нескольких видов прокариот. Процесс создания вновь синтезированной цепи из цепи-шаблона позволяет метилировать старую цепь в отдельное время от новой цепи. Это позволяет репаративным ферментам проверять новую цепь и исправлять любые мутации или ошибки.
ДНК может иметь способность активировать или деактивировать определенные области на вновь синтезированной цепи, что позволяет изменять фенотип клетки. Это может быть выгодно для клетки, потому что ДНК может активировать более благоприятный фенотип, чтобы помочь в выживании. Благодаря естественному отбору более благоприятный фенотип будет сохраняться у всего вида. Это порождает идею наследования или того, почему одни фенотипы наследуются над другими.
Источник
Биология. 11 класс
§ 16. Клеточный цикл. Репликация ДНК
Клеточный цикл . Все новые клетки образуются путем деления уже существующих. Этот принцип, как уже отмечалось, сформулировал Р. Вирхов еще в середине XIX в. Деление клеток обеспечивает непрерывность существования жизни на нашей планете. Именно благодаря ему осуществляются различные способы бесполого и полового размножения организмов. В основе процессов роста, регенерации и индивидуального развития многоклеточных организмов также лежит деление клеток.
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
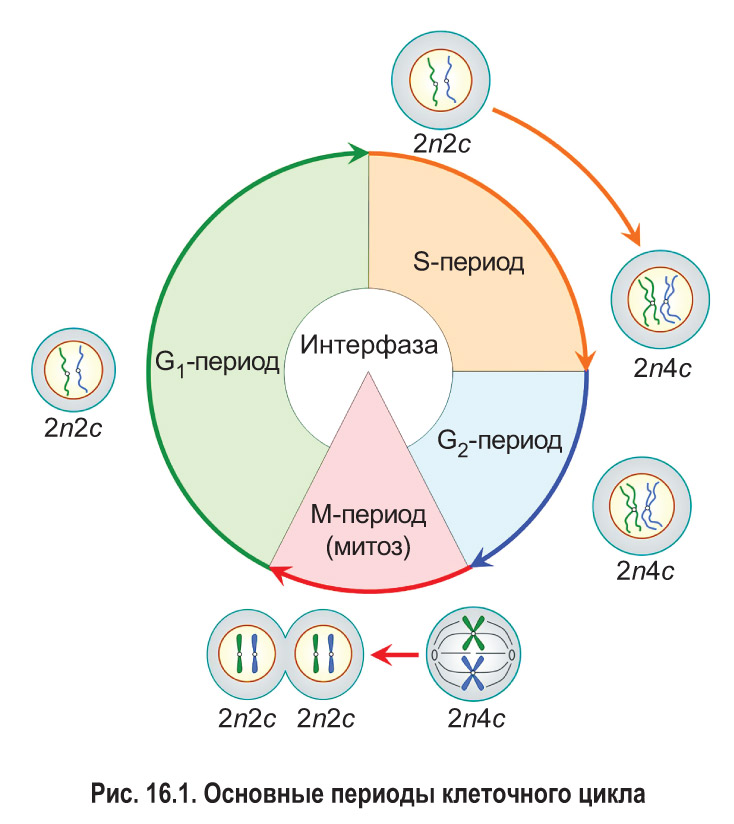 Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Итак, интерфаза обычно включает три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На протяжении всей интерфазы хромосомы не спирализованы. Они располагаются в ядре клетки в виде хроматина .
После завершения интерфазы начинается деление клетки. Основным способом деления клеток эукариот является митоз, поэтому данный этап клеточного цикла обозначают как М-период. При митозе происходит спирализация хроматина . Это приводит к формированию компактных двухроматидных хромосом. После этого сестринские хроматиды каждой хромосомы отделяются друг от друга и затем попадают в разные дочерние клетки. Значит, дочерние клетки, образовавшиеся в результате митоза и вступающие в новый клеточный цикл, имеют набор 2n2c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Источник
§ 16. Клеточный цикл. Репликация ДНК
| Сайт: | Профильное обучение |
| Курс: | Биология. 11 класс |
| Книга: | § 16. Клеточный цикл. Репликация ДНК |
| Напечатано:: | Гость |
| Дата: | Пятница, 19 Ноябрь 2021, 23:24 |
Оглавление
Клеточный цикл . Все новые клетки образуются путем деления уже существующих. Этот принцип, как уже отмечалось, сформулировал Р. Вирхов еще в середине XIX в. Деление клеток обеспечивает непрерывность существования жизни на нашей планете. Именно благодаря ему осуществляются различные способы бесполого и полового размножения организмов. В основе процессов роста, регенерации и индивидуального развития многоклеточных организмов также лежит деление клеток.
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Итак, интерфаза обычно включает три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На протяжении всей интерфазы хромосомы не спирализованы. Они располагаются в ядре клетки в виде хроматина .
После завершения интерфазы начинается деление клетки. Основным способом деления клеток эукариот является митоз, поэтому данный этап клеточного цикла обозначают как М-период. При митозе происходит спирализация хроматина . Это приводит к формированию компактных двухроматидных хромосом. После этого сестринские хроматиды каждой хромосомы отделяются друг от друга и затем попадают в разные дочерние клетки. Значит, дочерние клетки, образовавшиеся в результате митоза и вступающие в новый клеточный цикл, имеют набор 2n2c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Краткая характеристика
Пресинтети ческий (G1)
Рост клетки, образование органоидов, подготовка к репликации
Синтети ческий (S)
Репликация ДНК , удвоение центриолей клеточного центра
Постсинтети ческий (G2)
Завершение подготовки к делению
2n4c (в материнской клетке) → 2n2c (в каждой дочерней клетке)
Деление клетки на две дочерние
Некоторые клетки многоклеточных организмов, образовавшиеся при митозе, проходят далее G1-, S- и G2-периоды интерфазы и снова вступают в митоз. Это характерно, например, для клеток покровных эпителиев (росткового слоя эпидермиса кожи, эпителия желудочно-кишечного тракта и др.), красного костного мозга, образовательных тканей растений.
В отличие от них многие клетки после прохождения части G1-периода вступают в так называемый период покоя, или G0-период. Клетки, находящиеся в G0-периоде, выполняют свои функции в организме, однако в них не происходит подготовка к репликации. Это свойственно, прежде всего, высокоспециализированным клеткам — нейронам, клеткам сердечной мышцы, хрусталика глаза и др. Такие клетки, как правило, навсегда утрачивают способность к делению.
Однако некоторые клетки, пребывающие в G0-периоде (например, клетки печени, эндокринных желез, лейкоциты), сохраняют способность к выходу из периода покоя, продолжению клеточного цикла и последующему делению. Такое явление наблюдается, например, при повреждении органа, в состав которого данные клетки входят.
*Прохождение клетками определенных этапов клеточного цикла регулируется целым рядом механизмов. На протекание цикла влияют как внеклеточные сигналы (для одноклеточных организмов это действие факторов внешней среды, для многоклеточных — главным образом БАВ: гормоны, цитокины и др.), так и внутриклеточные. Важную роль в осуществлении контроля клеточного цикла играют белки циклины. Содержание тех или иных циклинов в клетках закономерно изменяется по мере прохождения определенных периодов цикла. Нарушение механизмов регуляции клеточного цикла может приводить к неконтролируемому размножению клеток, что является причиной образования опухолей.
В клеточном цикле существуют так называемые контрольные точки. Если клетка проходит такую точку, она продолжает «движение» по клеточному циклу. Если же какие-либо причины мешают клетке пройти через контрольную точку, то клеточный цикл останавливается. Следующей фазы цикла не наступает до устранения препятствий, которые не позволили клетке пройти контрольную точку.
Известны как минимум четыре контрольные точки клеточного цикла. Так, в конце G1-периода происходит проверка ДНК на наличие повреждений перед вступлением в S-период. При непрохождении этой точки, как правило, запускается процесс устранения повреждений — репарация ДНК (от лат. reparatio — восстановление). Следующий контроль осуществляется в конце S-периода — проверяется полнота репликации ДНК. В G2-периоде ДНК снова проверяется на наличие повреждений и завершенность репликации. Наконец в ходе митоза осуществляется контроль прикрепления ко всем хромосомам нитей веретена деления.*
Репликация ДНК . Вспомним, что удвоение молекул ДНК, которое осуществляется в синтетическом (S) периоде клеточного цикла, называется репликацией. *После открытия Дж. Уотсоном и Ф. Криком структуры молекулы ДНК было выдвинуто несколько гипотез о возможном механизме протекания этого процесса (рис. 16.2).
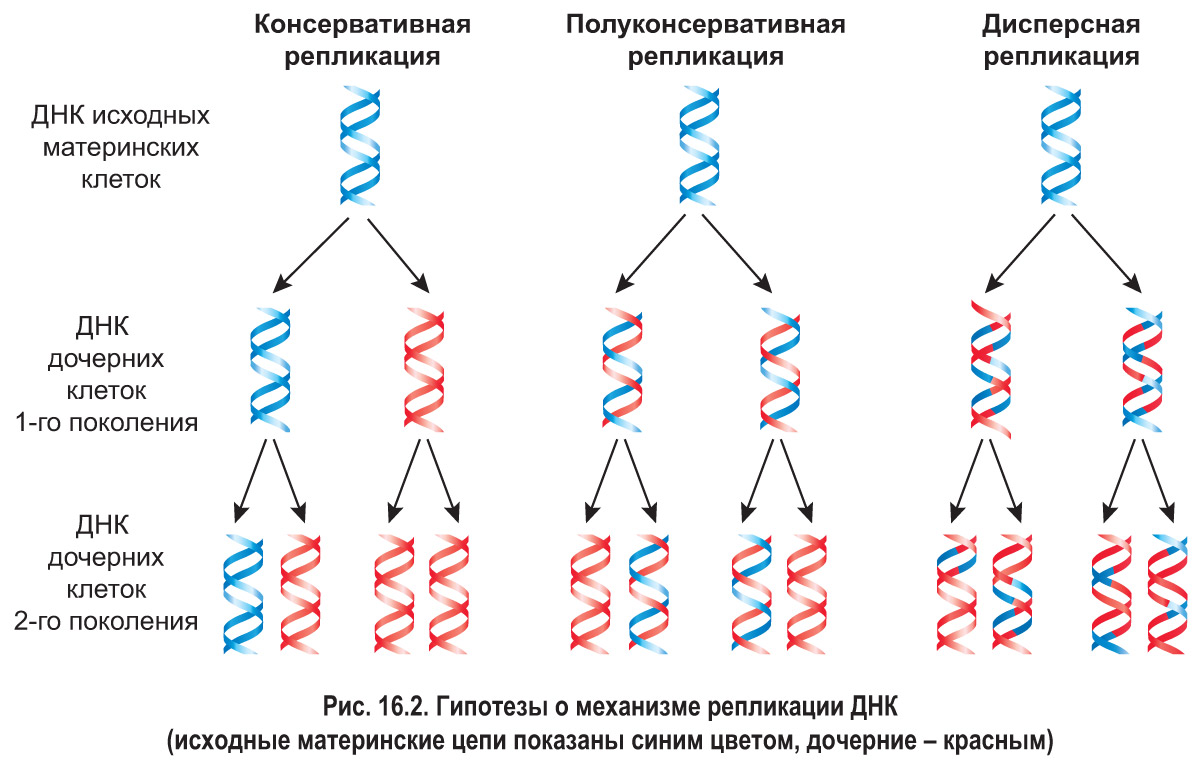
Согласно гипотезе консервативной репликации двухцепочечная материнская молекула ДНК как единое целое служит матрицей для образования дочерней молекулы, состоящей из двух совершенно новых цепей. Полуконсервативный механизм репликации предполагает разделение цепей исходной молекулы ДНК. При этом каждая материнская цепь является матрицей для синтеза дочерней цепи. Таким образом, при репликации образуются две молекулы ДНК, каждая из которых включает одну материнскую цепь и одну новую — дочернюю. Гипотеза дисперсной репликации состояла в том, что материнская ДНК распадается на фрагменты, которые выступают в роли матриц для построения отдельных участков новых молекул. Образованные таким способом молекулы ДНК должны состоять из чередующихся фрагментов исходной молекулы и вновь синтезированных.
В 1958 г. американские биологи М. Мезельсон и Ф. Сталь провели экспериментальную проверку этих гипотез. На протяжении нескольких поколений они выращивали бактерии (а именно кишечную палочку) в питательной среде, содержащей «тяжелый» азот 15 N. За это время 15 N вошел в состав бактериальных молекул ДНК. Далее бактерии были перенесены в среду, содержащую изотоп 14 N. Следовательно, в состав вновь синтезированных цепей ДНК включался уже «легкий» азот. Из бактериальных клеток новых поколений, образовавшихся в среде с 14 N, выделяли ДНК и центрифугировали в градиенте плотности хлорида цезия (рис. 16.3).

Выяснилось, что ДНК дочерних клеток первого поколения имела плотность, среднюю между плотностью «легкой» ДНК, содержащей только 14 N, и «тяжелой», включающей только 15 N. То есть такая «гибридная» ДНК содержала одновременно 14 N и 15 N. Это противоречило гипотезе о консервативном механизме репликации, согласно которой ДНК должна была разделиться на две фракции — «легкую» и «тяжелую».
ДНК, выделенная из клеток второго поколения, разделялась на «легкую» и «гибридную». Этот факт полностью соответствовал гипотезе полуконсервативной репликации и позволил исключить дисперсный механизм удвоения ДНК, согласно которому плотность ДНК второго поколения бактерий должна быть средней между «легкой» и «гибридной». Таким образом был доказан полуконсервативный механизм репликации ДНК.*
*К настоящему времени установлено, что в осуществлении репликации принимает участие целый комплекс ферментов. Также известно, что этот процесс начинается в строго определенных участках молекулы ДНК — так называемых точках начала репликации. Бактериальная хромосома, как правило, имеет одну такую точку. У ядерных организмов каждая молекула ДНК (хромосома) содержит множество точек начала репликации. Следовательно, у эукариот процесс репликации одновременно протекает на многих участках одной и той же хромосомы . Это значительно сокращает время удвоения молекул ДНК.
Процесс репликации ДНК подразделяют на три этапа: инициацию, элонгацию и терминацию.
1. Инициация (запуск). Особые ферменты начинают раскручивать молекулу ДНК от точки начала репликации. Затем фермент хеликаза разъединяет цепи исходной материнской молекулы, разрывая водородные связи между комплементарными азотистыми основаниями. При этом две цепи расходятся под определенным углом и образуют так называемую репликативную вилку, напоминающую букву Y (рис. 16.4). Хеликаза, подобно бегунку застежки «молния», движется вдоль молекулы ДНК, разделяя комплементарные цепи. Это сопровождается перемещением репликативной вилки и, образно говоря, расстегиванием «молнии». Далее с разошедшимися цепями связываются молекулы ДНК-полимеразы — главного фермента репликации.
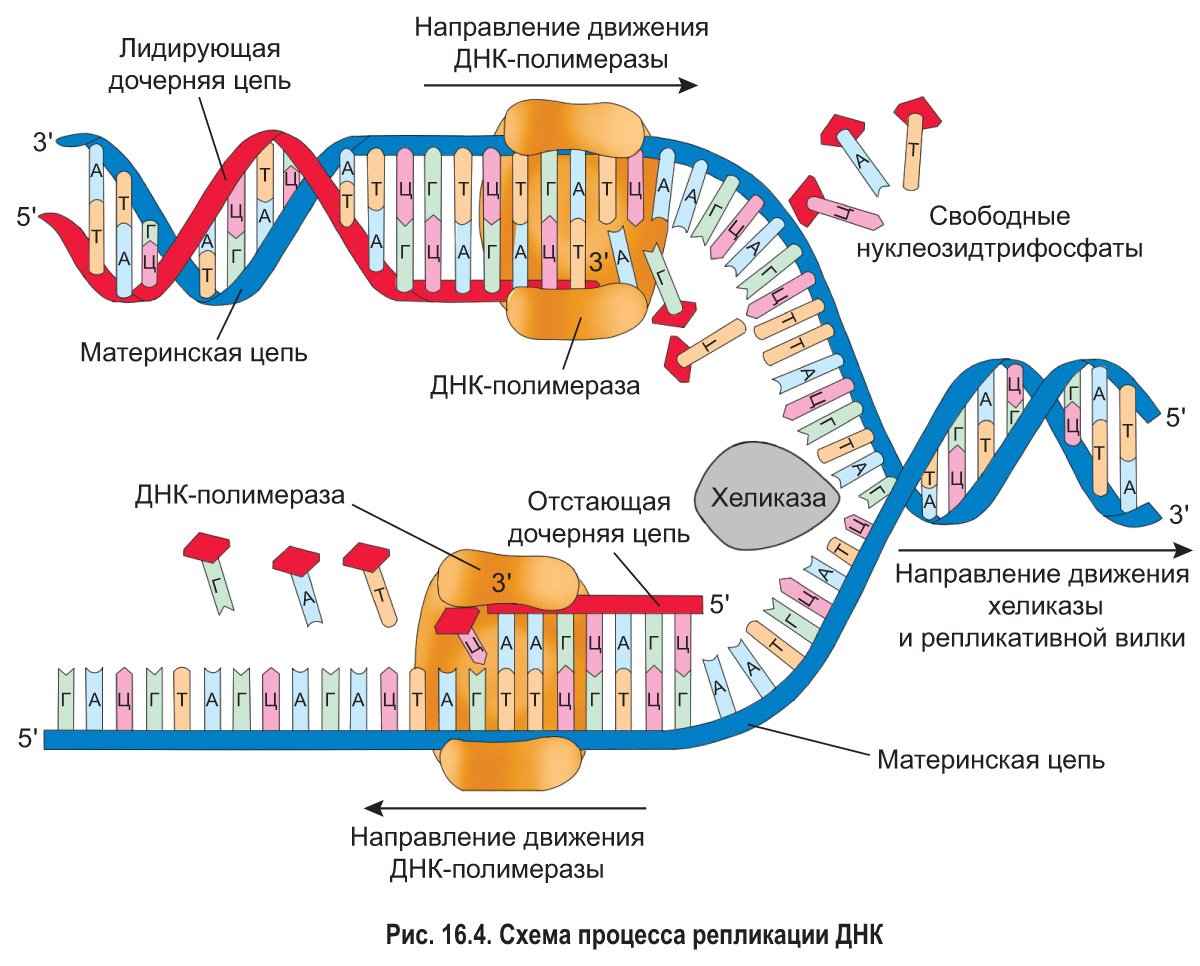
2. Элонгация (удлинение, наращивание дочерних цепей ДНК). Молекулы ДНК-полимеразы начинают двигаться вдоль материнских цепей, используя их в качестве матриц для построения новых дочерних цепей. Это значит, что в растущие дочерние цепи ДНК включаются только те нуклеотиды, которые комплементарны соответствующим нуклеотидам материнских цепей. Поэтому репликацию относят к реакциям матричного синтеза.
Следует отметить, что в процессе репликации материалом для синтеза дочерних цепей ДНК являются не нуклеотиды как таковые, а нуклеозидтрифосфаты. От обычных нуклеотидов их отличает наличие трех остатков фосфорной кислоты (как в молекуле АТФ) вместо одного. Связи между этими остатками являются макроэргическими. Таким образом, нуклеозидтрифосфаты — соединения, богатые энергией. В процессе их присоединения к растущим дочерним цепям ДНК происходит отщепление двух «лишних» остатков фосфорной кислоты. В результате выделяется энергия, которая используется для протекания реакции (т. е. для образования фосфоэфирных связей), и нуклеозидтрифосфаты становятся стандартными нуклеотидами новых цепей ДНК.
Особенность ДНК-полимеразы состоит в том, что она может двигаться вдоль существующей цепи ДНК только в направлении 3′ → 5′. При этом наращивание дочерней цепи всегда происходит антипараллельно: от 5′-конца к 3′-концу (т. е. новые нуклеотиды добавляются к 3′-концу синтезируемой цепи). Например, если участок материнской цепи ДНК содержит последовательность нуклеотидов 3’ГТАЦАГ5′, то при наращивании соответствующего ему участка дочерней цепи нуклеотиды будут присоединяться в следующем порядке: 5’ЦАТГТЦ3′.
Способность ДНК-полимеразы к перемещению вдоль цепи ДНК только в одном направлении приводит к следующему. Вдоль одной материнской цепи (той, у которой направление 3′ → 5′ совпадает с направлением перемещения репликативной вилки) фермент движется непрерывно. Дочерняя цепь ДНК, которая при этом синтезируется, называется лидирующей или ведущей (рис. 16.5).
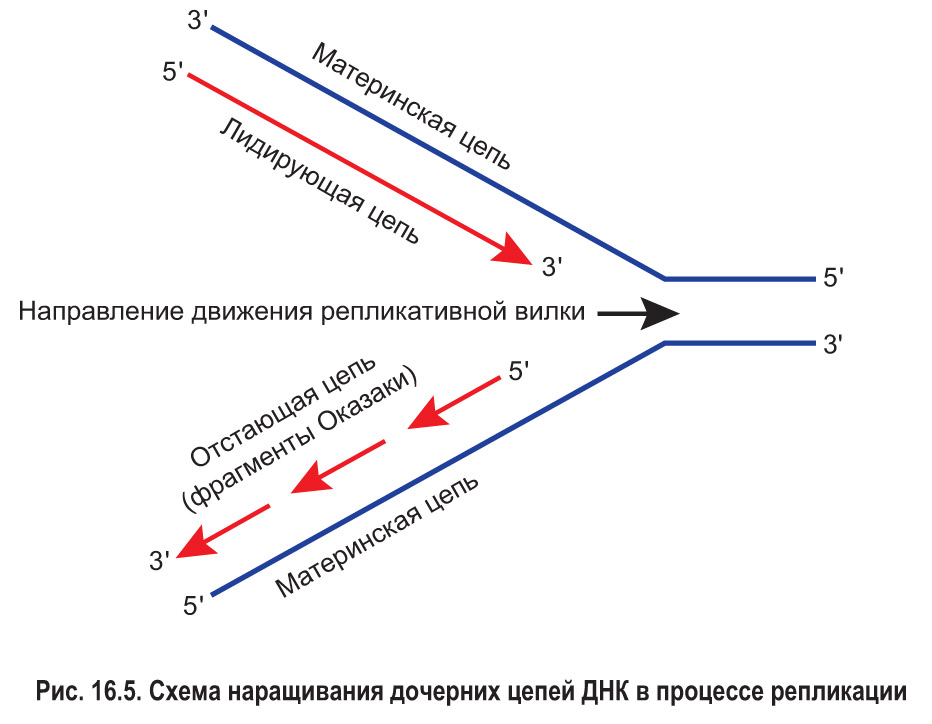
В то же время движение ДНК-полимеразы, которая работает над созданием другой дочерней цепи, не может быть непрерывным. По мере смещения репликативной вилки фермент «забегает вперед» и затем, возвращаясь назад, синтезирует новую цепь отдельными фрагментами. В честь первооткрывателей — супругов Р. и Ц. Оказáки — эти участки называют фрагментами Оказаки. Таким образом, вторая дочерняя цепь строится прерывисто с существенным отставанием от лидирующей цепи. Поэтому данная цепь получила название отстающей или запаздывающей. Впоследствии фрагменты отстающей цепи сшивает друг с другом фермент ДНК-лигаза.*
*Открытие фермента, катализирующего биосинтез ДНК, принадлежит американскому биохимику А. Корнбергу. Энзим, выделенный из клеток кишечной палочки, ученый назвал ДНК-полимеразой. В 1957 г. с помощью ДНК-полимеразы А. Корнберг впервые осуществил синтез ДНК в лабораторных условиях, а в 1959 г. за открытие механизмов биосинтеза нуклеиновых кислот он совместно с биохимиком С. Очоа был удостоен Нобелевской премии.*
*3. Терминация (остановка). Когда репликативная вилка достигает соседнего участка ДНК, на котором также осуществлялась репликация, ферменты завершают свою работу.* В результате репликации образуются две молекулы ДНК, идентичные друг другу и исходной материнской молекуле. В состав каждой из них входит одна цепь материнской молекулы ДНК и одна вновь синтезированная дочерняя цепь.
*Каждая клетка существует определенное время, по истечении которого либо делится, либо погибает. Различают два типа клеточной гибели — некроз и апоптоз.
Некроз — это гибель клеток, обусловленная действием повреждающих факторов. Причинами некроза может быть воздействие на клетки высоких или низких температур, ионизирующих излучений, токсических веществ, в том числе выделяемых болезнетворными микроорганизмами. Некротическая гибель клеток наблюдается также в результате их механического повреждения, нарушения иннервации и кровоснабжения тканей, в состав которых они входят, и т. д.
При некрозе обычно происходит гибель не отдельных клеток, а целых их групп. Например, в результате ожога массово погибают клетки кожи, подвергшиеся действию высокой температуры. При инфаркте миокарда из-за прекращения снабжения кислородом отмирает участок сердечной мышцы, состоящий из множества клеток. В поврежденных клетках нарушается проницаемость плазмалеммы и внутриклеточных мембран, прекращаются синтез белков и другие процессы метаболизма, происходит разрушение ядра и органоидов. Клетки погибают, и продукты их распада, как правило, стимулируют запуск воспалительной реакции в зоне некроза.
Апоптоз, который иногда называют клеточным самоубийством, представляет собой запрограммированную гибель клеток, которая регулируется организмом. Этот процесс обнаружен у различных групп живых организмов, в том числе и одноклеточных. У животных и растений апоптоз наблюдается на всех этапах индивидуального развития, даже в эмбриональный период. Известно, например, что в организме взрослого человека ежедневно в результате апоптоза запланированно погибает 50—70 млрд клеток.
Уничтожение клетки путем апоптоза могут запускать как определенные внеклеточные сигналы, так и внутриклеточные факторы. В результате апоптоза клетка распадается на отдельные фрагменты, окруженные плазмалеммой. Затем эти фрагменты поглощаются лейкоцитами или соседними клетками без запуска воспалительной реакции.
Благодаря апоптозу в организме поддерживается клеточный гомеостазис, т. е. определенное соотношение между различными типами клеток. Апоптоз играет важную роль в процессах клеточной дифференцировки, необходимой для правильного развития и функционирования тканей и органов. Он также обеспечивает уничтожение различных дефектных клеток, например мутантных. За открытие механизмов апоптоза и генетической регуляции развития органов британские биологи С. Бреннер и Дж. Салстон, а также американский ученый Р. Хорвиц в 2002 г. были удостоены Нобелевской премии.*

Период жизни клетки с момента ее образования из материнской клетки до деления (включая это деление) или гибели называется клеточным циклом. Он состоит из интерфазы и деления клетки. Интерфаза (часть клеточного цикла между делениями) обычно включает три периода. В G 1 -периоде клетка растет и готовится к репликации, в S-периоде происходит репликация, в G2-периоде завершается подготовка к делению. Многие клетки, вставшие на путь специализации, выходят из G 1 -периода в период покоя. Такие клетки не готовятся к репликации и в ряде случаев утрачивают способность к делению.
Репликация ДНК происходит с участием целого комплекса ферментов, важнейшим из которых является ДНК-полимераза. Матрицами для синтеза дочерних цепей ДНК служат обе цепи материнской молекулы, т. е. репликация является реакцией матричного синтеза. В результате репликации образуются две идентичные молекулы ДНК. В состав каждой из них входит одна цепь исходной материнской молекулы и одна вновь синтезированная дочерняя цепь. Таким образом, репликация осуществляется полуконсервативно.
 | 1. Что такое клеточный цикл? 2. Охарактеризуйте основные периоды клеточного цикла по плану: 1) название и сокращенное обозначение; 2) происходящие процессы; 3) содержание генетического материала в клетке. 3. Что представляет собой G0-период? Какие клетки в него вступают? Все ли клетки, вступающие в период покоя, навсегда утрачивают способность к делению? Ответ поясните. 4. Что такое репликация ДНК? Охарактеризуйте основные этапы репликации. 5. Почему репликацию ДНК относят к реакциям матричного синтеза? 6. В связи с чем при репликации синтез одной из дочерних цепей ДНК осуществляется непрерывно и сравнительно быстро, а другая цепь строится медленнее, отдельными фрагментами? 7*. Одинаковы ли молекулы ДНК, входящие в состав гомологичных хромосом? В состав сестринских хроматид? Почему? 8*. Опишите эксперимент, с помощью которого был доказан полуконсервативный механизм репликации ДНК. Как распределились бы молекулы ДНК в результате центрифугирования, если бы репликация протекала по консервативному механизму? Дисперсному? Пользуясь рисунками 16.2 и 16.3, нарисуйте соответствующие схемы. 9*. В многоклеточных организмах имеются так называемые стволовые клетки. Они являются первичными клетками, из которых в начале эмбрионального развития формируются органы зародыша, и на протяжении всей жизни сохраняют способность к делению. Раньше считалось, что стволовые клетки есть только у эмбрионов, теперь же они выявлены почти во всех органах взрослых людей. В 1999 г. журнал «Science» признал открытие стволовых клеток важнейшим научным событием в биологии. Каковы перспективы использования стволовых клеток? Источник |



