
Полуконсервативный способ репликации ДНК. Биологическое значение.
Полуконсервативный механизм репликации ДНК. Перед каждым делением клетки в ней должно удвоиться содержание ДНК, чтобы каждая дочерняя клетка получила полный набор хромосом. Основу каждой хромосомы образует одна двухцепочечная молекула ДНК. Предложенная Дж. Уотсоном и Ф. Криком модель строения ДНК форме регулярной двойной спирали сразу же позволила понять принцип копирования ДНК. Ее репликация происходит полуконсервативным способом: две исходные цепи материнской ДНК расходятся, и каждая из них становится матрицей для синтеза новой комплементарной цепи. Таким образом, каждая новая двойная спираль ДНК содержит одну старую и одну новую цепь. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух материнский цепей ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсон и Ф. Сталь.
Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся и каждая цепь служит матрицей для синтеза новой комплементарной цепи ДНК. Каждая дочерняя молекула состоит из одной старой материнской и одной новой синтезированной цепи ДНК.
Общие принципы репликации ДНК.
В основе процесса репликации лежит принцип копирования материнской цепи ДНК с образованием двух идентичных молекул ДНК. В основе синтеза новой цепи ДНК лежит принцип комплементарности азотистых оснований, т.е. последовательность нуклеотидов материнской цепи определяет последовательность нуклеотидов в синтезируемой цепи ДНК
Синтез новых цепей ДНК идет в направлении 5’ → 3’.
В основе репликации ДНК лежат принципы антипараллельности и униполярности. Синтез новых цепей ДНК идет в направлении от 5’-конца к 3’ — концу, при этом ДНК-полимеразы – ферменты, синтезирующие новые цепи ДНК, — добавляют нуклеотиды к 3’- концу наращиваемой цепи ДНК. При этом матричная цепь имеет противоположную ориентацию: 3’ → 5’, т.е. ДНК-полимеразы могут передвигаться вдоль матрицы только в направлении от 3’ к 5’-концам.
Механизмы синтеза ДНК хорошо изучены в клетках бактерий, однако имеются доказательства, что в клетках эукариот процесс протекает аналогичным образом.
Инициация репликации ДНК. Репликация начинается в специфическом участке молекулы ДНК, который называется точка начала репликации или ориджин.
Точка начала репликации (origin) – это участок молекулы ДНК со специфической последовательностью нуклеотидов с большим содержанием пар АТ (последовательность 300 п.н., богата АТ). Специальные инициирующие белки необходимы для связывания ферментов репликации с молекулой ДНК: белок DnaA – для прокариот белок RPA (replication protein A) – для эукриот.
Кольцевая хромосома прокариот имеет одну точку начала репликации, которая
называется OriC. В этой точке цепи расходятся и образуются две репликативные вилки, которые движутся в противоположном направлении. Скорость синтеза ДНК в клетках прокариот составляет 500 нукл./сек. Две вилки встречаются на противоположной стороне кольца. В клетках прокариот существует специальный фермент гираза (топоизомераза II), который разделяет две образующиеся кольцевые молекулы ДНК. Антибиотик налидиксовая кислота угнетает размножение бактерий, путем инактивации гиразы.
В клетках эукариот этот фермент отсутствует, поэтому налидиксовую кислоту используют в клинической практике для лечения бактериальных инфекций.
Репликация ДНК эукариот начинается одновременно во многих точках начала репликации, от каждой точки движутся две репликативные вилки в противоположных направлениях. Скорость синтеза молекул ДНК эукариот составляет 50 нукл./сек.
Репликон – фрагмент молекулы ДНК, репликация которого происходит под контролем одной точки начала репликации. Кольцевая хромосома прокариот имеет 1 репликон. Геномы эукариот содержат сотни и тысячи репликонов.
Инициация — образование репликативной вилки. Нити ДНК разделяются благодаря действию специальных ферментов и белков.
Хеликаза (от англ. helix – спираль) — основной фермент, расплетающий цепи ДНК. У прокариот он называется белок DnaB. Хеликаза разрывает водородные связи между комплементарными основаниями, используя энергию АТФ.
Топоизомеразы – ферменты, которые устраняют положительные сверхвитки перед репликативной вилкой. Эти ферменты временно разрывают нити материнской ДНК в двойной спирали перед репликативной вилкой, после завершения процесса нити ДНК восстанавливают целостность.
SSB — белки – это белки, которые связываются с одноцепочечной ДНК и удерживают матрицу. В результате образуется репликативная вилка, где и происходит синтез новых цепей ДНК.
Источник
Полуконсервативный способ репликации




![]()
![]()
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.
Гипотеза Уотсона-Крика была проверена с помощью опытов, выполненных М.Мезельсоном и Ф.Сталем в 1957 г. Клетки E.coli выращивали в течение ряда поколений в среде, содержащей в качестве источника азота хлористый аммоний NH4Cl, в котором обычный изотоп [ 14 N] был заменен на «тяжелый» изотоп [ 15 N]. Вследствие этого все соединения клеток, имеющие в своем составе азот, в том числе и азотистые основания ДНК, оказались обогащенными изотопом [ 15 N]. Плотность ДНК, выделенной из этих клеток, была выше плотности нормальной [ 14 N] ДНК. Смесь «тяжелой» [ 15 N] и «легкой» [ 14 N] ДНК удалось разделить методом центрифугирования в концентрированном растворе хлористого цезия. Поскольку [ 15 N] ДНК чуть тяжелее, чем [ 14 N] ДНК, полоса, в которой она достигает равновесия в градиенте СsCl, расположена ближе ко дну пробирки, чем полоса с [ 14 N] ДНК (рис.7)
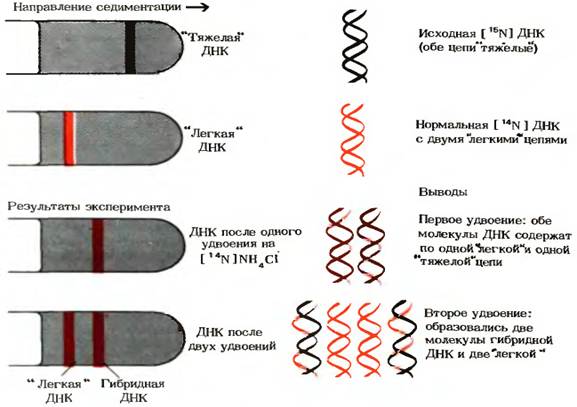
Рис. 7 Результаты эксперимента Мезельсона-Сталя
Мезельсон и Сталь перенесли клетки E.coli, росшие на среде с изотопом [ 15 N] и содержащие «тяжелые» цепи ДНК, на свежую среду с обычным изотопом [ 14 N]. На этой среде клетки E.coli выращивали в течение времени, необходимого для удвоения клеток. Затем из этих клеток выделяли ДНК и анализировали ее плотность с помощью описанного выше метода седиментации (осаждения ДНК в растворе СsCl). В градиенте СsCl была обнаружена лишь одна полоса ДНК, плотность которой оказалась средней между плотностью нормальной «легкой» [ 14 N] ДНК и плотностью «тяжелой» [ 15 N] ДНК (рис. ). Это подтвердило, что двухцепочечная ДНК дочерних клеток содержит одну старую 15 N – цепь от родительской ДНК и одну новую 14 N – цепь.
Если выделить ДНК из клеток, которые прошли два цикла удвоения на среде с [ 14 N], то она разделится на две полосы: одна с плотностью, соответствующей плотности нормальной «легкой» ДНК, а другая с плотностью «гибридной» ДНК, наблюдавшейся после первого удвоения клеток. На основании этих данных Мезельсон и Сталь пришли к выводу, что в строгом соответствии с гипотезой Уотсона-Крика каждый дочерний дуплекс ДНК после двух циклов удвоения клеток содержал одну родительскую и одну новообразованную цепь ДНК. Такой механизм репликации назвали полуконсервативным. Полученные результаты полностью исключили консервативный способ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсон и Сталя позволил также отвергнуть дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из коротких участков как родительской, так и новообразованнй ДНК, соединенных между собой случайным образом.
Источник
Полуконсервативный способ репликации днк биологическое значение

Biology exam запись закреплена
Экзаменационный вопрос №5.Способы репликации ДНК: консервативный, полуконсервативный, дисперсионный. Опыты Мезельсон и Сталь
В 1957 г. Метью Мезелсон и Франклин Сталь поставили один из самых красивых биологических экспериментов, исследуя механизмы репликации ДНК. Для определения способа репликации ДНК необходимо четко различать материнские и дочерние молекулы. Мезелсон и Сталь выращивали бактерии кишечной палочки (Escherichia coli) на среде, содержащей в качестве источника азота его тяжелый изотоп – 15N. Молекула ДНК содержит большое количество атомов азота, при делении ДНК азот для новых цепочек берется из внешней среды. Тяжелый изотоп азота включался в состав молекулы ДНК и служил надежной меткой. Для того чтобы пометить практически всю бактериальную ДНК, необходимо было культивировать E. coli на такой среде в течение как минимум 12 поколений.
Мезелсон и Сталь длительное время культивировали E. coli на среде с 15N. После этого бактерии быстро переносили на среду, содержащую более легкий изотоп азота 14N. Благодаря тому, что клетки E. coli культивировались на двух различных средах, в состав молекул их ДНК входили оба изотопа азота: 14N и 15N. Отличить такие молекулы друг от друга можно было по плотности, поскольку масса нуклеотидов молекулы ДНК, содержащих 15N больше, по сравнению с обычной молекулой. ДНК бактериальных клеток, выращенных на среде с 15N, имела плотность 1,724 г/см3, а ДНК клеток, выращенных на среде с 14N – 1,710 г/см3.
После переноса культуры E. coli с одной среды на другую через каждое поколение Мезелсон и Сталь отбирали пробы. Контролем служила бактериальная культура, содержавшаяся на среде с 15N. Из каждой пробы бактериальных клеток путем центрифугирования извлекали ДНК. После этого ее смешивали с раствором хлористого цезия (CsCl) плотностью 1,7 г/см3, и вновь центрифугировали с очень высокой скоростью в течение нескольких дней. В результате осаждения молекул CsCl его раствор приобретал градиент плотности от 1,65 г/см3 в верхней части пробирки до 1,8 г/см3 у ее дна. В соответствии с этим, молекулы ДНК концентрировались в строго определенной области: у дна, в центре или в верхней части пробирки. Локализация ДНК устанавливалась на спектрофотометре, поскольку было известно, что она поглощает лучи с длиной волны 260 нм. Этот метод Мезелсон и Сталь разработали и опубликовали совместно с Джермом Виноградом (1913–1976) в 1957 г., непосредственно перед проведением эксперимента по репликации ДНК. пределяя плотность ДНК в каждой из проб, Мезелсон и Сталь обнаружили, что спустя одно поколение после переноса культуры E. coli со среды с 15N на среду с 14N плотность ДНК была промежуточной между 15N-ДНК и 14N-ДНК. Спустя два поколения половина бактериальных клеток содержала ДНК с легким изотопом азота (14N), а другая половина – такую же, как и в предыдущем поколении, ДНК промежуточной плотности. Через три поколения на среде с 14N в ¾ клеток содержалась легкая ДНК, а ¼ часть клеток сохраняла ДНК промежуточной плотности. Т. о., соотношение между числом генераций и распределением плотности ДНК точно соответствовало полуконсервативному типу репликации.
месте с тем, из гипотезы полуконсервативной репликации следовало, что ДНК с промежуточной плотностью между 15N-ДНК и 14N-ДНК должна быть гибридной. Это значит, что одна из ее цепей должна содержать только тяжелый изотоп азота (15N), а другая – только легкий (14N). Проверяя это предположение, Мезелсон и Сталь нагревали полученную ими ДНК промежуточной плотности в течение 30 минут при температуре 100 °С на водяной бане. При таких условиях двойная спираль молекулы ДНК денатурирует, образуя две отдельных цепочки, однако ковалентные связи между нуклеотидами в каждой из них не разрушаются. Проведя центрифугирование денатурировавшей ДНК в градиенте плотности хлористого цезия, Мезелсон и Сталь обнаружили, что в результате образовалось две фракции различной плотности. Плотность одной из них совпадала с плотностью молекул ДНК, содержащих тяжелый изотоп азота (15N), тогда как плотность другой была идентична плотности молекул ДНК с легким изотопом азота (14N). Из этого следовало, что молекула ДНК промежуточной плотности, образовавшаяся в первом поколении после переноса со среды с тяжелым изотопом азота на среду с легким изотопом, представляет собой гибридную молекулу. В ее состав входят две цепи – материнская, содержащая исключительно 15N и вновь синтезированная дочерняя, содержащая только 14N. Эти данные также подтвердили верность модели Уотсона–Крика и полуконсервативный характер репликации ДНК.
— Консервативная репликация:
Молекула ДНК служит матрицей для образования совершенно новой молекулы ДНК. В результате одна из образующихся клеток получает исходную молекулу ДНК, а другая – вновь синтезированную.
— Полуконсервативная репликация:
Две цепи исходной молекулы ДНК расходятся вследствие разрыва слабых водородных связей между азотистыми основаниями. Каждая из них служит матрицей для образования новой цепи, а возникающие между азотистыми основаниями водородные связи соединяют старую и новую цепи, восстанавливая целостность молекулы. В результате каждая новая клетка получает гибридную молекулу ДНК, состоящую из одной старой и одной новой цепи.
— Дисперсная репликация:
ДНК распадается на короткие фрагменты, используемые в качестве матриц для построения фрагментов двух новых молекул ДНК, которые затем каким-то образом соединяются между собой.
Источник
Репликация днк Полуконсервативный механизм репликации
Для того чтобы объяснить, каким образом может самокопироваться, или редуплицироваться, такая стабильная и замкнутая на себя структура, как двойная спираль ДНК, Уотсон и Крик предположили, что ее цепи способны к раскручиванию и последующему частичному разделению вследствие разрыва водородных связей в каждой комплементарной паре оснований. Образовавшиеся одноцепочечные участки родительской молекулы могут служить матрицей, к которой на основе комплементарности оснований присоединяются соответствующие нуклеотиды. Эти нуклеотиды скрепляются между собой фосфодиэфирными связями с образованием новой цепи, комплементарной родительской. Так как этот процесс происходит на каждой разделившейся цепи исходной молекулы, то в результате образуются две двухцепочечные структуры, идентичные родительской ДНК. Такой способ репликации получил название полуконсервативного, поскольку в каждой из вновь образовавшейся молекул одна цепь является старой (родительской), а другая — вновь синтезированной (дочерней). Этот механизм обеспечивает возможность такого распределения ДНК между делящимися клетками, при котором каждая дочерняя клетка получает гибридную двухцепочечную молекулу ДНК, состоящую из родительской и вновь синтезированной цепи.
Первые данные в пользу гипотезы полуконсервативного механизма синтеза ДНК были получены Дж. Тейлором с соавторами (1957) цитологическим методом при изучении репликации хромосом конских бобов (Vicia faba). Экспериментально эта гипотеза была доказана с помощью физико-химических методов М. Мезельсоном и Ф. Сталем (1958). Сущность их метода заключалась в следующем: бактерии E.coli на протяжении многих генераций выращивали в среде, содержащей в качестве источника азота только его тяжелый изотоп 15 N. Эта метка включалась в азотсодержащие пуриновые и пиримидиновые основания в ДНК, вследствие чего ДНК в клетках, выращенных среде с 15 N, имела большую молекулярную массу на единицу объема (т.е. большую плотность), чем ДНК в клетках, выращенных в обычных условиях, в присутствии легкого изотопа 14 N. Поэтому, если клетки после длительного выращивания на среде с 15 N отмывали и переносили на время, равное одной, двум и т.д. генерациям, в среду с 14 N вместо 15 N, то это должно было привести к появлению молекул ДНК с меньшей плотностью. Различающиеся по плотности молекулы можно разделить с помощью метода равновесного ультрацентрифугирования в градиенте плотности 6М CsCl. Оказалось, что вся ДНК, выделенная из клеток, выращенных в течение одной генерации в среде с 14 N, располагается в градиенте CsCl в положении, промежуточном между положением “тяжелой” ДНК из клеток, выращенных только в среде с 15 N, и “легкой” ДНК из клеток, выращенных только в среде с 14 N. Это значит, что по прошествии одной генерации после переноса из среды с 15 N в среду с 14 N, в клетках появилась гибридная по плотности ДНК, у которой одна цепь была “тяжелой”, а другая — “легкой”. Если ДНК выделяли из клетки через две генерации после такого переноса, то “гибридная” ДНК составляла лишь половину всей ДНК, распределяющейся в градиенте плотности CsCl, а другая половина располагалась в зоне, соответствующей по плотности молекулам, включавшим только 14 N. В следующем поколении эта “легкая” фракция увеличивалась и составляла 75% тотальной ДНК. Такое изменение можно объяснить только с позиций представления о полуконсервативном способе репликации ДНК.
Сделанный Мезельсоном и Сталем вывод был полностью подтвержден и для других объектов, включая животных и высшие растения. Вместе с тем, оставалось неясным, в каком направлении происходит репликация ДНК и какие ферменты обеспечивают этот процесс. Ответ на первый вопрос удалось получить Дж. Кэрнсу (1963) в опытах на E.coli. Он поставил своей задачей сделать процесс репликации ДНК видимым, для чего использовал метод авторадиографии. Этот метод позволяет обнаруживать и локализовывать радиоактивную метку, включающуюся в молекулы ДНК, РНК, либо белка путем их помещения на чувствительную к радиации фотоэмульсию. ДНК можно специфично метить, если выращивать клетки с 3 Н-тимидином, являющимся дезоксирибонуклеозидом тимина. Тимидин избирательно включается в ДНК, поэтому выращивание клеток E.coli в среде с меченым по тритию тимидином позволяет следить за распределением метки по мере ее включения во вновь синтезирующуюся ДНК. Поскольку крупные молекулы ДНК легко разрываются при механических повреждениях, Кэрнс разработал такую технику, при которой клетки разрушались ферментом лизоцимом, а ДНК из лизированных клеток собирали на мембранных фильтрах, помещаемых на стекла, покрытые фотоэмульсией, чувствительной к -частицам, выделяющимся при распаде трития. Радиоавтографы, полученные после проявления таких фотографических стекол, выдерживавшихся предварительно в темноте до образования зерен металлического серебра, показали, что хромосома E.coli имеет кольцевидную форму и во время своей репликации образует структуры в форме греческой буквы . Обнаружение подобных структур, являющихся формами молекул ДНК в процессе их репликации, показывало, что раскручивание двух комплементарных цепей родительской ДНК и их полуконсервативная репликация происходят практически одновременно и начинаются в общей точке начала репликации, обозначаемой как локус ori (от англ. origin- начало). Последующий анализ результатов опыта Кэрнса и данные других экспериментаторов, нацеленных на изучение механизма репликации ДНК у различных бактерий, фагов, плазмид, показали, что в большинстве случаев она происходит двунаправлено и начинается, как правило, от одного уникального локуса ori. В этом месте в одной из цепей ДНК разрывается фосфодиэфирная связь, обеспечивающая последующее раскручивание дуплекса и образование особых структур — репликативных вилок, движущихся в противоположных направлениях по кольцевой ДНК. Следует, однако, отметить, что в ДНК эукариот, как правило, обнаруживается не один, а множество локусов ori, что, по-видимому, служит необходимым условием для того, чтобы громадные молекулы ДНК в хромосомах эукариот успели полностью отреплицироваться за время одного клеточного цикла.
Эксперименты, проведенные на прокариотах (E.coli, T7, некоторые плазмиды), также выявили в их ДНК (скорость репликации которых составляет около 20 мкм/мин, что в 20-30 раз выше, чем у эукариот) не по одному, а по два или даже более локусов ori. Среди этих локусов наблюдается подчиненность: когда активен главный ori, остальные могут не функционировать, и, напротив, в случае выключения главного ori запасные ori обеспечивают репликацию ДНК. Очевидно, такая зависимость обеспечивает стабильность важнейшей функции ДНК — ее способности к репликации.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Источник



