
Полуконсервативный способ репликации ДНК. Биологическое значение.
Полуконсервативный механизм репликации ДНК. Перед каждым делением клетки в ней должно удвоиться содержание ДНК, чтобы каждая дочерняя клетка получила полный набор хромосом. Основу каждой хромосомы образует одна двухцепочечная молекула ДНК. Предложенная Дж. Уотсоном и Ф. Криком модель строения ДНК форме регулярной двойной спирали сразу же позволила понять принцип копирования ДНК. Ее репликация происходит полуконсервативным способом: две исходные цепи материнской ДНК расходятся, и каждая из них становится матрицей для синтеза новой комплементарной цепи. Таким образом, каждая новая двойная спираль ДНК содержит одну старую и одну новую цепь. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух материнский цепей ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсон и Ф. Сталь.
Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся и каждая цепь служит матрицей для синтеза новой комплементарной цепи ДНК. Каждая дочерняя молекула состоит из одной старой материнской и одной новой синтезированной цепи ДНК.
Общие принципы репликации ДНК.
В основе процесса репликации лежит принцип копирования материнской цепи ДНК с образованием двух идентичных молекул ДНК. В основе синтеза новой цепи ДНК лежит принцип комплементарности азотистых оснований, т.е. последовательность нуклеотидов материнской цепи определяет последовательность нуклеотидов в синтезируемой цепи ДНК
Синтез новых цепей ДНК идет в направлении 5’ → 3’.
В основе репликации ДНК лежат принципы антипараллельности и униполярности. Синтез новых цепей ДНК идет в направлении от 5’-конца к 3’ — концу, при этом ДНК-полимеразы – ферменты, синтезирующие новые цепи ДНК, — добавляют нуклеотиды к 3’- концу наращиваемой цепи ДНК. При этом матричная цепь имеет противоположную ориентацию: 3’ → 5’, т.е. ДНК-полимеразы могут передвигаться вдоль матрицы только в направлении от 3’ к 5’-концам.
Механизмы синтеза ДНК хорошо изучены в клетках бактерий, однако имеются доказательства, что в клетках эукариот процесс протекает аналогичным образом.
Инициация репликации ДНК. Репликация начинается в специфическом участке молекулы ДНК, который называется точка начала репликации или ориджин.
Точка начала репликации (origin) – это участок молекулы ДНК со специфической последовательностью нуклеотидов с большим содержанием пар АТ (последовательность 300 п.н., богата АТ). Специальные инициирующие белки необходимы для связывания ферментов репликации с молекулой ДНК: белок DnaA – для прокариот белок RPA (replication protein A) – для эукриот.
Кольцевая хромосома прокариот имеет одну точку начала репликации, которая
называется OriC. В этой точке цепи расходятся и образуются две репликативные вилки, которые движутся в противоположном направлении. Скорость синтеза ДНК в клетках прокариот составляет 500 нукл./сек. Две вилки встречаются на противоположной стороне кольца. В клетках прокариот существует специальный фермент гираза (топоизомераза II), который разделяет две образующиеся кольцевые молекулы ДНК. Антибиотик налидиксовая кислота угнетает размножение бактерий, путем инактивации гиразы.
В клетках эукариот этот фермент отсутствует, поэтому налидиксовую кислоту используют в клинической практике для лечения бактериальных инфекций.
Репликация ДНК эукариот начинается одновременно во многих точках начала репликации, от каждой точки движутся две репликативные вилки в противоположных направлениях. Скорость синтеза молекул ДНК эукариот составляет 50 нукл./сек.
Репликон – фрагмент молекулы ДНК, репликация которого происходит под контролем одной точки начала репликации. Кольцевая хромосома прокариот имеет 1 репликон. Геномы эукариот содержат сотни и тысячи репликонов.
Инициация — образование репликативной вилки. Нити ДНК разделяются благодаря действию специальных ферментов и белков.
Хеликаза (от англ. helix – спираль) — основной фермент, расплетающий цепи ДНК. У прокариот он называется белок DnaB. Хеликаза разрывает водородные связи между комплементарными основаниями, используя энергию АТФ.
Топоизомеразы – ферменты, которые устраняют положительные сверхвитки перед репликативной вилкой. Эти ферменты временно разрывают нити материнской ДНК в двойной спирали перед репликативной вилкой, после завершения процесса нити ДНК восстанавливают целостность.
SSB — белки – это белки, которые связываются с одноцепочечной ДНК и удерживают матрицу. В результате образуется репликативная вилка, где и происходит синтез новых цепей ДНК.
Источник
полуконсервативная репликация
Смотреть что такое «полуконсервативная репликация» в других словарях:
полуконсервативная репликация — Метод репликации молекулы ДНК, при которой каждая материнская цепь молекулы ДНК достраивает себе новую дочернюю цепь [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN semiconservative replication … Справочник технического переводчика
Полуконсервативная репликация — * паўкансерватыўная рэплікацыя * semiconservative replication тип репликации ДНК, при котором молекула делится продольно, каждая половина сохраняется и служит матрицей для образующейся новой нити. Термин был введен, когда точный процесс… … Генетика. Энциклопедический словарь
Репликация полуконсервативная — * рэплікацыя паўкансерватыўная * semi conservative replication процесс репликации, при котором дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь синтезируется вновь. Такой же процесс повторяется при образовании… … Генетика. Энциклопедический словарь
репликация θ-типа — репликация θ типа θ репликация Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования «вздутия», видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Справочник технического переводчика
θ-репликация — θ type replication репликация θ типа, θ репликация. Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования вздутия , видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Молекулярная биология и генетика. Толковый словарь.
Семиконсервативная репликация — * семікансерватыўная рэплікацыя * semiconservative replication … Генетика. Энциклопедический словарь
генетический код — Долгое время наследственные механизмы были предметом внимания генетиков, но природа молекул, переносящих информацию от одного индивида к другому, оставалась неведомой. Была известна роль макромолекул протеина и нуклеиновых кислот в этом… … Западная философия от истоков до наших дней
История науки — История науки это исследование феномена науки в его истории. Наука, в частности, представляет собой совокупность эмпирических, теоретических и практических знаний о Мире, полученных научным сообществом. Поскольку с одной стороны наука… … Википедия
semiconservative replication — semiconservative replication. См. полуконсервативная репликация. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
5 -гидроксиметилцитозин 5 -гМц — 5 гидроксиметилцитозин, 5 гМц * 5 гідроксіметылцытазін * 5 hydroxymethyl cytosine or 5 hMC пиримидиновое основание, замещающее цитозин в ДНК четных колифагов. 5 ГМЦ комплементарен гуанину. Считается, что фагоспецифичная ДНКаза бактериофага… … Генетика. Энциклопедический словарь
Источник
Полуконсервативного способа репликации ДНК
Экспериментальное доказательство
Удвоение (репликация) ДНК
Принцип комплементарности (дополнения), лежащий в основе структуры ДНК, позволяет понять, как синтезируются новые молекулы ДНК незадолго перед делением клетки. Этот синтез обусловлен замечательной способностью молекулы ДНК к удвоению, он же определяет передачу наследственных свойств от материнской клетки к дочерним.
Сборка новой цепи идет в точном соответствии с принципом комплементарности: против А-Т, против Г-Ц, в результате образуется две молекулы ДНК того же нуклеотидного состава. Одна цепь «старая» происходит от первоначальной, а другая – синтезируется вновь. Таким образом, воспроизводится точная копия «материнской» молекулы ДНК. Иными словами, каждая нить ДНК служит матрицей, на которой строится комплементарная ей цепь. Процесс удвоения ДНК называется матричным синтезом, т.к. новые молекулы ДНК имеют ту же структуру, что и исходящие.
В дочерних клетках сохраняется та же наследственная информация. Если в исходной ДНК произойдет перестановка или замена нуклеотидов, то в дочерних это будет скопировано – в этом заключается молекулярный механизм изменчивости. Это изменение будет передаваться из клетки в клетку, из поколения в поколение.
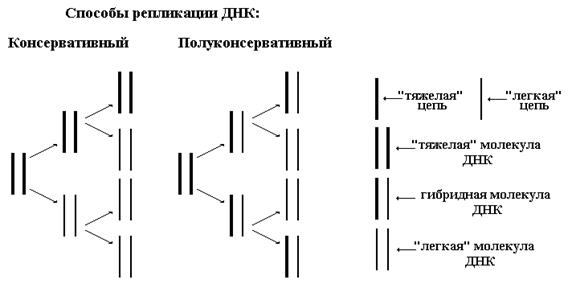
До получения экспериментального подтверждения полуконсервативного способа репликации предлагались три варианта удвоения ДНК:
Н рисунке ниже изображены схемы консервативного и полуконсервативного способов репликации ДНК.

Мозаичный способ предполагает деградацию материнской ДНК до олигонуклеотидных фрагментов и дальнейшую сборку двух новых молекул ДНК как из этих фрагментов, так и с использованием новых нуклеотидов из цитоплазмы (кариоплазмы). Данная гипотеза сразу же вызвала большое количество нареканий, так как требует наличия аппарата запоминания информации, хранящейся в материнской ДНК, на время ее разрушения.
Таким образом, консервативный способ предполагает, что материнская ДНК полностью переходит к одной из дочерних клеток, полуконсервативный — каждой из дочерних клеток переходит по одной цепи от материнской ДНК, мозаичный — обе новые ДНК построены из смеси старых и новых олигонуклеотидных фрагментов.
В 1958 году Месельсон и Сталь с помощью только что разработанного метода центрифугирования в градиенте плотности доказали, что молекулы ДНК удваиваются полуконсервативным способом. Для этого E. coli выращивали в течение многих поколений в среде, содержащей в качестве единственного источника азота 15 NH4Cl. В результате такой процедуры весь азот 14 N, содержащийся в исходной ДНК замещался на “тяжелый” 15 N. Молекулы 14 N-ДНК и 15 N-ДНК различаются по массе всего на 1%. Однако этой разницы достаточно для разделения таких молекул ДНК в градиенте плотности хлористого цезия. Формирование линейного градиента плотности в центрифужной пробирке осуществляется при длительном центрифугировании пробирок, содержащих возрастающие сверху до низу концентрации хлористого цезия. В результате создается устойчивый градиент плотности от 1,66 до 1,76 г/см 3 . Под действием центробежной силы молекулы ДНК переходят в ту область градиента, в которой плотность раствора равна их собственной плавучей плотности. Высокополимерные молекулы ДНК образуют четко выраженную полосу, которую легко обнаружить по поглощению в ультрафиолете.
После полного замещения 14 N в молекулах ДНК на “тяжелый” 15 N, бактериальные клетки переносили в среду, содержащую 14 NH4Cl. При этом в результате последовательных клеточных циклов, каждый из которых включает репликацию ДНК, происходило постепенное обратное замещение «тяжелых» цепей ДНК на «легкие». Динамика изменения «тяжести» молекул ДНК при этом свидетельствовала о способе репликации. На рисунке показаны 3 возможных варианта размещения молекул ДНК в центрифужной пробирке в зависимости от их «тяжести».
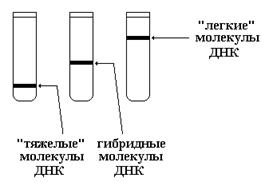
При консервативной и полуконсервативном способах репликации должны были наблюдаться следующие изменения положения полос ДНК в центрифужной пробирке после каждого клеточного цикла.

Таким образом, полуконсервативный способ репликации ДНК, характеризуется исчезновением после первого же деления клеток «тяжелых» молекул ДНК, а также появлением гибридных молекул.
Мозаичный способ репликации характеризоваться появлением после первого деления расплывчатой полосы в районе гибридных ДНК с постепенным ее сдвигом вверх при последующих делениях.
Результаты, полученные Месельсоном и Сталем, свидетельствовали именно о полуконсерватив-ном способе репликации ДНК.
Источник
Полуконсервативного способа репликации ДНК
До получения экспериментального подтверждения полуконсервативного способа репликации предлагались три варианта удвоения ДНК:
Н рисунке ниже изображены схемы консервативного и полуконсервативного способов репликации ДНК.
Мозаичный способ предполагает деградацию материнской ДНК до олигонуклеотидных фрагментов и дальнейшую сборку двух новых молекул ДНК как из этих фрагментов, так и с использованием новых нуклеотидов из цитоплазмы (кариоплазмы). Данная гипотеза сразу же вызвала большое количество нареканий, так как требует наличия аппарата запоминания информации, хранящейся в материнской ДНК, на время ее разрушения.
Таким образом, консервативный способ предполагает, что материнская ДНК полностью переходит к одной из дочерних клеток, полуконсервативный — каждой из дочерних клеток переходит по одной цепи от материнской ДНК, мозаичный — обе новые ДНК построены из смеси старых и новых олигонуклеотидных фрагментов.
В 1958 году Месельсон и Сталь с помощью только что разработанного метода центрифугирования в градиенте плотности доказали, что молекулы ДНК удваиваются полуконсервативным способом. Для этого E. coli выращивали в течение многих поколений в среде, содержащей в качестве единственного источника азота 15 NH4Cl. В результате такой процедуры весь азот 14 N, содержащийся в исходной ДНК замещался на “тяжелый” 15 N. Молекулы 14 N-ДНК и 15 N-ДНК различаются по массе всего на 1%. Однако этой разницы достаточно для разделения таких молекул ДНК в градиенте плотности хлористого цезия. Формирование линейного градиента плотности в центрифужной пробирке осуществляется при длительном центрифугировании пробирок, содержащих возрастающие сверху до низу концентрации хлористого цезия. В результате создается устойчивый градиент плотности от 1,66 до 1,76 г/см 3 . Под действием центробежной силы молекулы ДНК переходят в ту область градиента, в которой плотность раствора равна их собственной плавучей плотности. Высокополимерные молекулы ДНК образуют четко выраженную полосу, которую легко обнаружить по поглощению в ультрафиолете.
После полного замещения 14 N в молекулах ДНК на “тяжелый” 15 N, бактериальные клетки переносили в среду, содержащую 14 NH4Cl. При этом в результате последовательных клеточных циклов, каждый из которых включает репликацию ДНК, происходило постепенное обратное замещение «тяжелых» цепей ДНК на «легкие». Динамика изменения «тяжести» молекул ДНК при этом свидетельствовала о способе репликации. На рисунке показаны 3 возможных варианта размещения молекул ДНК в центрифужной пробирке в зависимости от их «тяжести».
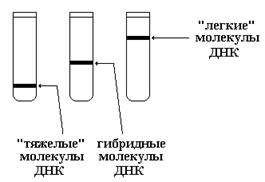
При консервативной и полуконсервативном способах репликации должны были наблюдаться следующие изменения положения полос ДНК в центрифужной пробирке после каждого клеточного цикла.

Таким образом, полуконсервативный способ репликации ДНК, характеризуется исчезновением после первого же деления клеток «тяжелых» молекул ДНК, а также появлением гибридных молекул.
Мозаичный способ репликации характеризоваться появлением после первого деления расплывчатой полосы в районе гибридных ДНК с постепенным ее сдвигом вверх при последующих делениях.
Результаты, полученные Месельсоном и Сталем, свидетельствовали именно о полуконсерватив-ном способе репликации ДНК.
Источник



