
Полуконсервативный способ репликации ДНК. Биологическое значение.
Полуконсервативный механизм репликации ДНК. Перед каждым делением клетки в ней должно удвоиться содержание ДНК, чтобы каждая дочерняя клетка получила полный набор хромосом. Основу каждой хромосомы образует одна двухцепочечная молекула ДНК. Предложенная Дж. Уотсоном и Ф. Криком модель строения ДНК форме регулярной двойной спирали сразу же позволила понять принцип копирования ДНК. Ее репликация происходит полуконсервативным способом: две исходные цепи материнской ДНК расходятся, и каждая из них становится матрицей для синтеза новой комплементарной цепи. Таким образом, каждая новая двойная спираль ДНК содержит одну старую и одну новую цепь. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух материнский цепей ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсон и Ф. Сталь.
Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся и каждая цепь служит матрицей для синтеза новой комплементарной цепи ДНК. Каждая дочерняя молекула состоит из одной старой материнской и одной новой синтезированной цепи ДНК.
Общие принципы репликации ДНК.
В основе процесса репликации лежит принцип копирования материнской цепи ДНК с образованием двух идентичных молекул ДНК. В основе синтеза новой цепи ДНК лежит принцип комплементарности азотистых оснований, т.е. последовательность нуклеотидов материнской цепи определяет последовательность нуклеотидов в синтезируемой цепи ДНК
Синтез новых цепей ДНК идет в направлении 5’ → 3’.
В основе репликации ДНК лежат принципы антипараллельности и униполярности. Синтез новых цепей ДНК идет в направлении от 5’-конца к 3’ — концу, при этом ДНК-полимеразы – ферменты, синтезирующие новые цепи ДНК, — добавляют нуклеотиды к 3’- концу наращиваемой цепи ДНК. При этом матричная цепь имеет противоположную ориентацию: 3’ → 5’, т.е. ДНК-полимеразы могут передвигаться вдоль матрицы только в направлении от 3’ к 5’-концам.
Механизмы синтеза ДНК хорошо изучены в клетках бактерий, однако имеются доказательства, что в клетках эукариот процесс протекает аналогичным образом.
Инициация репликации ДНК. Репликация начинается в специфическом участке молекулы ДНК, который называется точка начала репликации или ориджин.
Точка начала репликации (origin) – это участок молекулы ДНК со специфической последовательностью нуклеотидов с большим содержанием пар АТ (последовательность 300 п.н., богата АТ). Специальные инициирующие белки необходимы для связывания ферментов репликации с молекулой ДНК: белок DnaA – для прокариот белок RPA (replication protein A) – для эукриот.
Кольцевая хромосома прокариот имеет одну точку начала репликации, которая
называется OriC. В этой точке цепи расходятся и образуются две репликативные вилки, которые движутся в противоположном направлении. Скорость синтеза ДНК в клетках прокариот составляет 500 нукл./сек. Две вилки встречаются на противоположной стороне кольца. В клетках прокариот существует специальный фермент гираза (топоизомераза II), который разделяет две образующиеся кольцевые молекулы ДНК. Антибиотик налидиксовая кислота угнетает размножение бактерий, путем инактивации гиразы.
В клетках эукариот этот фермент отсутствует, поэтому налидиксовую кислоту используют в клинической практике для лечения бактериальных инфекций.
Репликация ДНК эукариот начинается одновременно во многих точках начала репликации, от каждой точки движутся две репликативные вилки в противоположных направлениях. Скорость синтеза молекул ДНК эукариот составляет 50 нукл./сек.
Репликон – фрагмент молекулы ДНК, репликация которого происходит под контролем одной точки начала репликации. Кольцевая хромосома прокариот имеет 1 репликон. Геномы эукариот содержат сотни и тысячи репликонов.
Инициация — образование репликативной вилки. Нити ДНК разделяются благодаря действию специальных ферментов и белков.
Хеликаза (от англ. helix – спираль) — основной фермент, расплетающий цепи ДНК. У прокариот он называется белок DnaB. Хеликаза разрывает водородные связи между комплементарными основаниями, используя энергию АТФ.
Топоизомеразы – ферменты, которые устраняют положительные сверхвитки перед репликативной вилкой. Эти ферменты временно разрывают нити материнской ДНК в двойной спирали перед репликативной вилкой, после завершения процесса нити ДНК восстанавливают целостность.
SSB — белки – это белки, которые связываются с одноцепочечной ДНК и удерживают матрицу. В результате образуется репликативная вилка, где и происходит синтез новых цепей ДНК.
Источник
Полуконсервативным способом как это

Biology exam запись закреплена
Экзаменационный вопрос №5.Способы репликации ДНК: консервативный, полуконсервативный, дисперсионный. Опыты Мезельсон и Сталь
В 1957 г. Метью Мезелсон и Франклин Сталь поставили один из самых красивых биологических экспериментов, исследуя механизмы репликации ДНК. Для определения способа репликации ДНК необходимо четко различать материнские и дочерние молекулы. Мезелсон и Сталь выращивали бактерии кишечной палочки (Escherichia coli) на среде, содержащей в качестве источника азота его тяжелый изотоп – 15N. Молекула ДНК содержит большое количество атомов азота, при делении ДНК азот для новых цепочек берется из внешней среды. Тяжелый изотоп азота включался в состав молекулы ДНК и служил надежной меткой. Для того чтобы пометить практически всю бактериальную ДНК, необходимо было культивировать E. coli на такой среде в течение как минимум 12 поколений.
Мезелсон и Сталь длительное время культивировали E. coli на среде с 15N. После этого бактерии быстро переносили на среду, содержащую более легкий изотоп азота 14N. Благодаря тому, что клетки E. coli культивировались на двух различных средах, в состав молекул их ДНК входили оба изотопа азота: 14N и 15N. Отличить такие молекулы друг от друга можно было по плотности, поскольку масса нуклеотидов молекулы ДНК, содержащих 15N больше, по сравнению с обычной молекулой. ДНК бактериальных клеток, выращенных на среде с 15N, имела плотность 1,724 г/см3, а ДНК клеток, выращенных на среде с 14N – 1,710 г/см3.
После переноса культуры E. coli с одной среды на другую через каждое поколение Мезелсон и Сталь отбирали пробы. Контролем служила бактериальная культура, содержавшаяся на среде с 15N. Из каждой пробы бактериальных клеток путем центрифугирования извлекали ДНК. После этого ее смешивали с раствором хлористого цезия (CsCl) плотностью 1,7 г/см3, и вновь центрифугировали с очень высокой скоростью в течение нескольких дней. В результате осаждения молекул CsCl его раствор приобретал градиент плотности от 1,65 г/см3 в верхней части пробирки до 1,8 г/см3 у ее дна. В соответствии с этим, молекулы ДНК концентрировались в строго определенной области: у дна, в центре или в верхней части пробирки. Локализация ДНК устанавливалась на спектрофотометре, поскольку было известно, что она поглощает лучи с длиной волны 260 нм. Этот метод Мезелсон и Сталь разработали и опубликовали совместно с Джермом Виноградом (1913–1976) в 1957 г., непосредственно перед проведением эксперимента по репликации ДНК. пределяя плотность ДНК в каждой из проб, Мезелсон и Сталь обнаружили, что спустя одно поколение после переноса культуры E. coli со среды с 15N на среду с 14N плотность ДНК была промежуточной между 15N-ДНК и 14N-ДНК. Спустя два поколения половина бактериальных клеток содержала ДНК с легким изотопом азота (14N), а другая половина – такую же, как и в предыдущем поколении, ДНК промежуточной плотности. Через три поколения на среде с 14N в ¾ клеток содержалась легкая ДНК, а ¼ часть клеток сохраняла ДНК промежуточной плотности. Т. о., соотношение между числом генераций и распределением плотности ДНК точно соответствовало полуконсервативному типу репликации.
месте с тем, из гипотезы полуконсервативной репликации следовало, что ДНК с промежуточной плотностью между 15N-ДНК и 14N-ДНК должна быть гибридной. Это значит, что одна из ее цепей должна содержать только тяжелый изотоп азота (15N), а другая – только легкий (14N). Проверяя это предположение, Мезелсон и Сталь нагревали полученную ими ДНК промежуточной плотности в течение 30 минут при температуре 100 °С на водяной бане. При таких условиях двойная спираль молекулы ДНК денатурирует, образуя две отдельных цепочки, однако ковалентные связи между нуклеотидами в каждой из них не разрушаются. Проведя центрифугирование денатурировавшей ДНК в градиенте плотности хлористого цезия, Мезелсон и Сталь обнаружили, что в результате образовалось две фракции различной плотности. Плотность одной из них совпадала с плотностью молекул ДНК, содержащих тяжелый изотоп азота (15N), тогда как плотность другой была идентична плотности молекул ДНК с легким изотопом азота (14N). Из этого следовало, что молекула ДНК промежуточной плотности, образовавшаяся в первом поколении после переноса со среды с тяжелым изотопом азота на среду с легким изотопом, представляет собой гибридную молекулу. В ее состав входят две цепи – материнская, содержащая исключительно 15N и вновь синтезированная дочерняя, содержащая только 14N. Эти данные также подтвердили верность модели Уотсона–Крика и полуконсервативный характер репликации ДНК.
— Консервативная репликация:
Молекула ДНК служит матрицей для образования совершенно новой молекулы ДНК. В результате одна из образующихся клеток получает исходную молекулу ДНК, а другая – вновь синтезированную.
— Полуконсервативная репликация:
Две цепи исходной молекулы ДНК расходятся вследствие разрыва слабых водородных связей между азотистыми основаниями. Каждая из них служит матрицей для образования новой цепи, а возникающие между азотистыми основаниями водородные связи соединяют старую и новую цепи, восстанавливая целостность молекулы. В результате каждая новая клетка получает гибридную молекулу ДНК, состоящую из одной старой и одной новой цепи.
— Дисперсная репликация:
ДНК распадается на короткие фрагменты, используемые в качестве матриц для построения фрагментов двух новых молекул ДНК, которые затем каким-то образом соединяются между собой.
Источник
Полуконсервативного способа репликации ДНК
До получения экспериментального подтверждения полуконсервативного способа репликации предлагались три варианта удвоения ДНК:
Н рисунке ниже изображены схемы консервативного и полуконсервативного способов репликации ДНК.
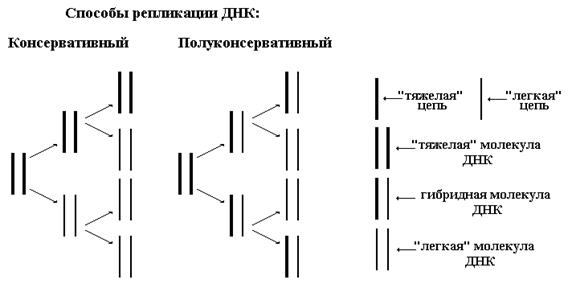
Мозаичный способ предполагает деградацию материнской ДНК до олигонуклеотидных фрагментов и дальнейшую сборку двух новых молекул ДНК как из этих фрагментов, так и с использованием новых нуклеотидов из цитоплазмы (кариоплазмы). Данная гипотеза сразу же вызвала большое количество нареканий, так как требует наличия аппарата запоминания информации, хранящейся в материнской ДНК, на время ее разрушения.
Таким образом, консервативный способ предполагает, что материнская ДНК полностью переходит к одной из дочерних клеток, полуконсервативный — каждой из дочерних клеток переходит по одной цепи от материнской ДНК, мозаичный — обе новые ДНК построены из смеси старых и новых олигонуклеотидных фрагментов.
В 1958 году Месельсон и Сталь с помощью только что разработанного метода центрифугирования в градиенте плотности доказали, что молекулы ДНК удваиваются полуконсервативным способом. Для этого E. coli выращивали в течение многих поколений в среде, содержащей в качестве единственного источника азота 15 NH4Cl. В результате такой процедуры весь азот 14 N, содержащийся в исходной ДНК замещался на “тяжелый” 15 N. Молекулы 14 N-ДНК и 15 N-ДНК различаются по массе всего на 1%. Однако этой разницы достаточно для разделения таких молекул ДНК в градиенте плотности хлористого цезия. Формирование линейного градиента плотности в центрифужной пробирке осуществляется при длительном центрифугировании пробирок, содержащих возрастающие сверху до низу концентрации хлористого цезия. В результате создается устойчивый градиент плотности от 1,66 до 1,76 г/см 3 . Под действием центробежной силы молекулы ДНК переходят в ту область градиента, в которой плотность раствора равна их собственной плавучей плотности. Высокополимерные молекулы ДНК образуют четко выраженную полосу, которую легко обнаружить по поглощению в ультрафиолете.
После полного замещения 14 N в молекулах ДНК на “тяжелый” 15 N, бактериальные клетки переносили в среду, содержащую 14 NH4Cl. При этом в результате последовательных клеточных циклов, каждый из которых включает репликацию ДНК, происходило постепенное обратное замещение «тяжелых» цепей ДНК на «легкие». Динамика изменения «тяжести» молекул ДНК при этом свидетельствовала о способе репликации. На рисунке показаны 3 возможных варианта размещения молекул ДНК в центрифужной пробирке в зависимости от их «тяжести».
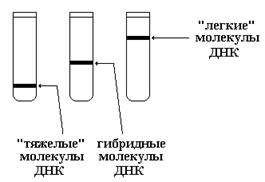
При консервативной и полуконсервативном способах репликации должны были наблюдаться следующие изменения положения полос ДНК в центрифужной пробирке после каждого клеточного цикла.

Таким образом, полуконсервативный способ репликации ДНК, характеризуется исчезновением после первого же деления клеток «тяжелых» молекул ДНК, а также появлением гибридных молекул.
Мозаичный способ репликации характеризоваться появлением после первого деления расплывчатой полосы в районе гибридных ДНК с постепенным ее сдвигом вверх при последующих делениях.
Результаты, полученные Месельсоном и Сталем, свидетельствовали именно о полуконсерватив-ном способе репликации ДНК.
Источник
полуконсервативная репликация
Смотреть что такое «полуконсервативная репликация» в других словарях:
полуконсервативная репликация — Метод репликации молекулы ДНК, при которой каждая материнская цепь молекулы ДНК достраивает себе новую дочернюю цепь [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN semiconservative replication … Справочник технического переводчика
Полуконсервативная репликация — * паўкансерватыўная рэплікацыя * semiconservative replication тип репликации ДНК, при котором молекула делится продольно, каждая половина сохраняется и служит матрицей для образующейся новой нити. Термин был введен, когда точный процесс… … Генетика. Энциклопедический словарь
Репликация полуконсервативная — * рэплікацыя паўкансерватыўная * semi conservative replication процесс репликации, при котором дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь синтезируется вновь. Такой же процесс повторяется при образовании… … Генетика. Энциклопедический словарь
репликация θ-типа — репликация θ типа θ репликация Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования «вздутия», видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Справочник технического переводчика
θ-репликация — θ type replication репликация θ типа, θ репликация. Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования вздутия , видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Молекулярная биология и генетика. Толковый словарь.
Семиконсервативная репликация — * семікансерватыўная рэплікацыя * semiconservative replication … Генетика. Энциклопедический словарь
генетический код — Долгое время наследственные механизмы были предметом внимания генетиков, но природа молекул, переносящих информацию от одного индивида к другому, оставалась неведомой. Была известна роль макромолекул протеина и нуклеиновых кислот в этом… … Западная философия от истоков до наших дней
История науки — История науки это исследование феномена науки в его истории. Наука, в частности, представляет собой совокупность эмпирических, теоретических и практических знаний о Мире, полученных научным сообществом. Поскольку с одной стороны наука… … Википедия
semiconservative replication — semiconservative replication. См. полуконсервативная репликация. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
5 -гидроксиметилцитозин 5 -гМц — 5 гидроксиметилцитозин, 5 гМц * 5 гідроксіметылцытазін * 5 hydroxymethyl cytosine or 5 hMC пиримидиновое основание, замещающее цитозин в ДНК четных колифагов. 5 ГМЦ комплементарен гуанину. Считается, что фагоспецифичная ДНКаза бактериофага… … Генетика. Энциклопедический словарь
Источник



