
Типы питания прокариот. Группы микроорганизмов по типу питания
В отличие от растительных и животных организмов, имеющих один вполне определенный тип питания – соответственно автотрофный и гетеротрофный, прокариоты характеризуются многообразием типов питания. Поэтому для характеристики типов питания трокариотных организмов используются одновременно три критерия: источник углерода, источник энергии и донор электронов (водорода).
По источнику углерода прокариоты являются автотрофами, если они получают углерод в результате фиксации углекислого газа, и гетеротрофами, если источником углерода для них служат органические соединения.
По источнику энергии прокариоты, использующие солнечный свет, называются фототрофами, а получающие энергию за счет окислительно-восстановительных реакций – хемотрофами.
И наконец, по донору электронов прокариоты подразделяются на литотрофы, обладающие способностью использовать неорганические вещества, органотрофы, использующие в качестве доноров электрона органические соединения.
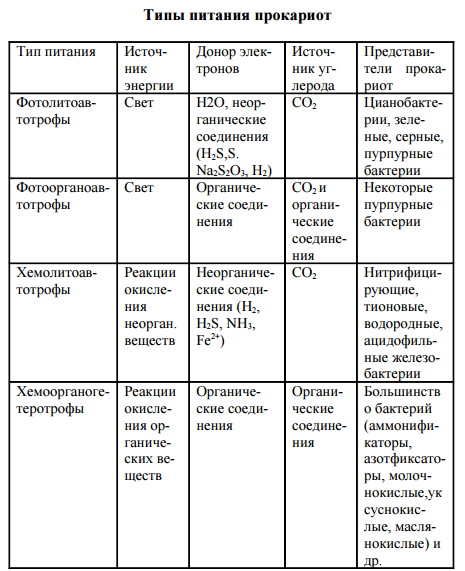
По трем вышеуказанным критериям выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы хемолитоавтотрофы и хемоорганогетеротрофы (таблица 1).
Различают следующие типы гетеротрофии: паразитизм облигатный внутриклеточный, паразитизм факультативный, сапрофитизм.
Организмы, которые могут жить только внутри других организмов, имеют редуцированный метаболизм, зависят от метаболизма хозяина, являются облигатными внутриклеточными паразитами.
Факультативные паразиты-организмы, способные расти вне клетки хозяина при подходящих условиях, на питательных средах.
Сапрофиты – гетеротрофные организмы, нуждаются в готовых органических веществах, непосредственно от организмов не зависят.
Сапрофиты нуждаются в разных концентрациях органических веществ. Олиготрофы – способны расти при низких концентрациях органического вещества (1–15 мг углерода в литре раствора). Копиотрофы – предпочитают высокие концентрации питательных веществ (10 грамм углерода в литре раствора).
Дата добавления: 2015-08-14 ; просмотров: 4896 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Прокариоты и эукариоты — что это и в чем их отличия
Здравствуйте, уважаемые читатели блога KtoNaNovenkogo.ru.
Все живущие на Земле организмы в зависимости от структуры их клеток относятся к одной из двух групп: прокариоты или эукариоты.

Термины «прокариоты» и «эукариоты» были предложены французским ботаником Э.Шаттоном в 1925 г.
Деление организмов на прокариотические и эукариотические сохранялось довольно долго (до 1990-х гг.), пока американский микробиолог К.Вёзе не обнаружил, что в среде прокариотов находится большая группа особей с существенными генетическими различиями.
В этой связи он предложил разделить прокариотов на бактерии и археи. В настоящий момент разделение живых организмов на эукариотов, бактерии и археи считается общепризнанным.
Прокариоты — это.
Прокариоты – это одноклеточные живые организмы без оформленного клеточного ядра. Они не развиваются, не переходят в многоклеточную форму и способны к автономному существованию.
Прокариоты – самая представительная форма жизни на Земле по количеству видов. Например, 1 грамм плодородной почвы может содержать порядка 10 млрд.бактериальных клеток.
Как уже отмечено выше, к прокариотам относятся бактерии (в том числе цианобактерии или сине-зелёные водоросли) и археи.
У прокариотов молекула органического вещества не отделена от цитоплазмы, а прикреплена к клеточной мембране. У них, как правило, бесполый способ размножения, а ДНК имеет кольцевую форму. У большинства прокариотов геном (что это?) представлен одиночной хромосомой.

Прокариоты – это древнейшие и в то же время самые примитивные организмы на нашей планете. Они встречаются повсеместно: в воздухе, в воде, в почве, внутри живых организмов.
Их можно обнаружить в океанических глубинах, на горных вершинах, во льдах Антарктиды и Арктики. В атмосфере споры бактерий присутствуют на высоте до 15 км, а в грунт они проникают на глубину более 4 км.
По форме бактериальные клетки отличаются огромным разнообразием. Они могут быть в виде палочек (бациллы), округлыми (диплококи), шестиугольными, звездообразными, стебельковыми и т.д. Диплококки образуют пары, стрептококки – цепочки, стафилококки – скопления наподобие виноградных гроздей.
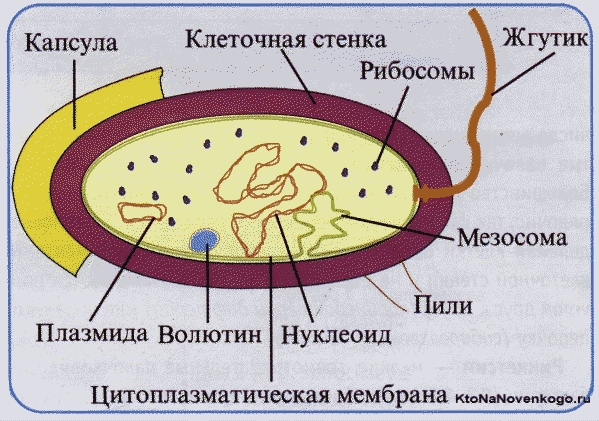
Строение бактериальной клетки в упрощённом виде выглядит следующим образом:
- клеточная оболочка (стенка);
- плазматическая мембрана;
- цитоплазма;
- хромосомная кольцевая ДНК (прикреплена к мембране);
- плазмиды (небольшие не прикреплённые к мембране кольцевые ДНК с небольшим набором генов);
- рибосомы;
- прокариотический жгутик(и).
Подавляющее большинство прокариот размножается посредством простого бинарного деления, которое начинается с удвоения ДНК без образования хромосом.
Обе вновь образовавшиеся молекулы ДНК отделяются друг от друга плазматической мембраной, в результате чего клетка делится пополам. Таким образом, каждая дочерняя клетка содержит по одной равнозначной молекуле ДНК.
Процесс деления при благоприятных условиях происходит каждые 25-30 минут. Этот интервал может увеличиться под воздействием сдерживающих факторов, таких как нехватка пищи, солнечный свет, высокая температура и др.
По способу питания бактерии делятся на гетеротрофов (это как?) и автотрофов (это как?).
Первые представлены сапротрофами (питаются мёртвой органикой), паразитами (потребляют органику живых особей) и симбионтами (живут и питаются вмести с другими организмами). Вторые получают питание посредством фотосинтеза (путём преобразования солнечной энергии либо за счёт химического окисления неорганических веществ).
Эукариоты — это.
В отличие от прокариотов, эукариоты – это ядерные живые организмы (т.е. их клетки содержат ядро).
Они могут быть как одноклеточными, так и многоклеточными, однако строение клеток у них однотипное.
В группу эукариотов (они могут быть одно- или многоклеточными) входят растения, животные (в том числе человек) и грибы.
Клетки эукариот разделены системой мембран на отдельные отсеки, имеют схожий химический состав и однотипный обмен веществ.
Генетический материал сконцентрирован, главным образом, в хромосомах, которые образованы цепочками ДНК и белковыми молекулами. В цитоплазме располагаются мембранные органоиды.

Непременным структурным элементом любой эукариотической клетки является ядро. В нём, а также в митохондриях животные клетки хранят наследственную информацию.
В растительных клетках эта информация находится не только в ядре и митохондриях, но ещё и в пластидах. Объёмное соотношение между ядром и цитоплазмой называется ядерно-цитоплазматическим индексом, с помощью которого можно оценить уровень метаболизма (это что?).
Почему грибы принадлежат к группе эукариот
У клеток грибов есть оформленное ядро, поэтому их относят к эукариотам.
Правда, изначально к эукариотам относили только растения и животных. В дальнейшем были выделены грибы как отдельное царство, так как они сочетают в себе растительные и животные признаки.
В частности, у них отсутствует хлорофилл, а питание происходит путём впитывания органических веществ из внешней среды (создавать собственную органику они не способны). Размножаются грибы как половым, так и бесполым способом.
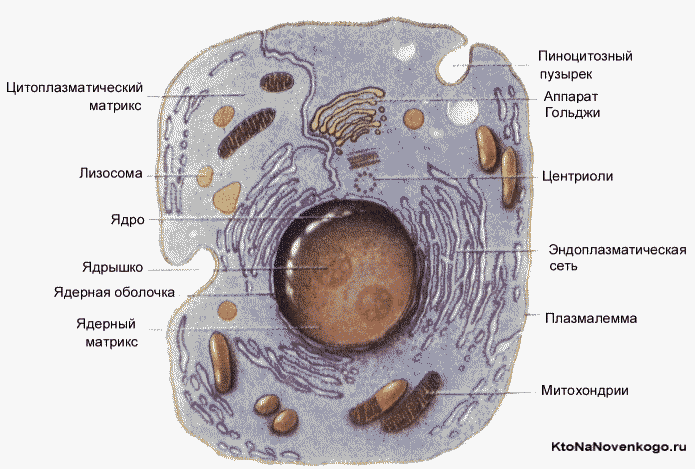
В состав клетки эукариот входят следующие основные компоненты:
- ядро;
- ядерная мембрана;
- линейная ДНК;
- цитоплазма;
- митохондрии;
- плазматическая или клеточная мембрана;
- хромосомы;
- рибосомы;
- лизосомы (у животных клеток для переваривания клеточных микромолекул);
- хлоропласты (у растительных клеток для обеспечения фотосинтеза);
- эукариотический жгутик(и).
Согласно самым распространённым научным гипотезам эукариоты появились порядка 1,5 млрд.лет тому назад. Многие учёные полагают, что они эволюционировали благодаря симбиогенезу, т.е. взаимодействию собственных клеток с клетками бактерий.
Отличие прокариотов от эукариотов
Главное, что отличает прокариотов от эукариотов, – отсутствие клеточного ядра.
А это значит, что ДНК прокариотической клетки не организована в хромосомы и не окружена ядерной оболочкой. Эукариотические клетки устроены намного сложнее. Их ДНК упакована в хромосомы, которые располагаются как раз в ядре.
Основные отличия рассматриваемых биологических категорий сведены в таблицу:
| Прокариоты | Эукариоты |
|---|---|
| Одноклеточные (за редким исключением) | Одно- или многоклеточные |
| Не имеют сформировавшегося ядра | Имеют чётко выраженное ядро (ядра) с собственной оболочкой |
| Наследственная информация содержится в кольцевой молекуле ДНК | Наследственная информация хранится в линейной ДНК ядра, а также митохондриях и пластидах |
| Не имеют мембранных органоидов | Содержат мембранные органоиды и немембранные структуры |
| Бинарное деление клетки | Прямое деление (амитоз), непрямое деление (митоз) или редукционное деление (мейоз) |
| Набор генов – гаплоидный | Набор генов, как правило, – диплоидный |
| Размножение вегетативное, споровое, почкованием | Размножение половое с образованием гамет |
| Жгутик в виде белковых нитей вмонтирован в оболочку клетки | Жгутик представлен выростом клетки в виде микротрубки |
| Клетки имеют размер 0,1-10 мкм | Клетки имеют размер 10-100 мкм |
» alt=»»>
Удачи вам! До скорых встреч на страницах блога KtoNaNovenkogo.ru
Эта статья относится к рубрикам:
Комментарии и отзывы (4)
Происхождение эукариотов является одной из самых больших тайн эволюции. Ученые до сих пор не знают, как возникло ядро и главное — почему? Есть мнение, что для качественного перехода прокариотам понадобилось войти в контакт с некими клетками или бактериями, которые либо уже существовали на планете, либо попали на Землю извне. В результате их симбиоза и возникли эукариоты.
Неприятно думать и вообще задумываться на эту тему, что нас окружают миллиарды микроскопических организмов. Хорошо, что глаз их никак не улавливает, иначе можно было бы свихнуться. А вообще, природа загадочна и удивительна!
На Марс недавно сел очередной планетоход, как раз в русло древней высохшей реки, было бы здорово, если бы этот марсоход обнаружил
там древних марсианских бактерий, пусть и мёртвых, всё равно это была бы сенсация.
Цианобактерии производят метан, а он в свою очередь усугубляет ситуацию с глобальным потеплением, поскольку этот газ создаёт парниковый эффект.
Источник
Типы питания прокариот
Прокариоты характеризуются многообразием типов питания в отличие от растений (автотрофы) и животных (ге- теротрофы). Для характеристики типов питания используют три критерия: источник углерода, источник энергии, донор электронов (водорода). По источнику углерода делят на ав- тотрофы и гетеротрофы, по источнику энергии – на фото- трофы (используют солнечный свет) и хемотрофы (получа- ют энергию за счет окислительно-восстановительных реак- ций), по донору – электроны (водорода) на литотрофы (неорганические соединения Н2, NH3, H2S, Fe2 + , CO и др.) и органотрофы (используют в качестве донора электрона ор- ганические соединения). Таким образом, выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы, хемолитоавтотрофы, хемоорганоге- теротрофы.
Фотолитоавтотрофы. Бактериальный фотосинтез (фоторедукция). В 1931 г. К. ван Ниль впервые доказал способность бактерий к фотосинтезу. В основе бактериального фотосинтеза лежит превращение световой энергии, поглощаемой фотосинтетическим пигментом, в биохимическую энергию макроэргических связей (АТФ) и далее использо- вание этой энергии для восстановления углекислого газа в процессе биосинтеза. У этих организмов есть пигмент бак- териохлорофилл. В клетках всех фотосинтезирующих бактерий содержатся фотосинтетические пигменты. К ним относятся особые хлорофиллы, получившие название бактериохлорофиллов а, в, с, d, и каротиноиды. По строению бактерио- хлорофиллы близки к хлорофиллу а растений. Так, бакте- риохлорофилл а отличается от хлорофилла а растений тем, что в первом его пирольном кольце в положении 2 стоит ацетильная группа СН3-СО- вместо обычной винильной группы СН2=СН-, а второе пирольное кольцо восстановле- но и содержит на два атома водорода больше. Содержание в клетке фотосинтезирующих прокариот четырех видоизменений бактериохлорофилла обусловли- вает более широкую полосу спектра поглощения световой энергии по сравнению с хлорофиллом растений. Предел спектра поглощения хлорофилла растений лежит в области 700-780 нм, в то время как фотосинтезирующие бактерии используют световую энергию длинноволновой части спектра до 1100 нм. Разница в спектрах поглощения хлоро- филлов растений и бактериохлорофиллов фотосинтезиру- ющих бактерий расширяет экологические ниши последних и позволяет фотосинтезирующим бактериям развиваться в водоемах под слоем водорослей. Помимо бактериохлорофиллов, в клетках фотосинтезирующих бактерий открыты более 20 дополнительных каро- тиноидных пигментов. Особенно разнообразны каротинои- ды в клетках пурпурных бактерий. Каротиноиды фотосин- тезирующих бактерий поглощают световую энергию ко- ротковолновой видимой области спектра с длиной волны 400-550 нм и передают эту энергию на бактериохлоро- филл. В клетке прокариот фотосинтетические пигменты нахо- дятся на инвагинациях ЦПМ – хроматофорах, визикулах (пузырьках), трубочках, тиллакоидах, ламеллах. Ван Ниль назвал этот процесс бактериальным фотосин- тезом, или фоторедукцией.
Для восстановления одной молекулы углекислого газа требуется 1 молекула АТФ (у растений 4 АТФ). В том и другом случае идет восстановление углекислого газа до углеводов. В Определителе Берджи (1974) фотосинтезирующие бактерии представлены тремя семействами (см. выше).
Фотоорганоавтотрофы Фотоорганоавтотрофы представлены немногочисленным семейством Rhodospirillaceae, включающим три рода: Rhodospirillum, Rhodopseudomonas и Rhodomicrobium. Бактерии-фотоорганоавтотрофы способны перестраивать свой обмен и одинаково успешно развиваться как на свету, так и в темноте, переходя соответственно от ана- эробного образа жизни к аэробному. На свету они ведут себя как фотоорганоавтотрофы: усваивая углекислый газ, они восстанавливают его в процессе фотосинтеза до углевода. В качестве доноров электронов несерные пурпурные бактерии используют различные органические вещества — сахара, спирты, органические кислоты, аминокислоты. Характер использования органических веществ различными фотосинтезирующими бактериями существенно раз- личается. Чаще всего органическое вещество выполняет единственную функцию донора электронов при фотоасси- миляции углекислого газа. Однако в некоторых случаях органическое вещество используется фотосинтезирующими бактериями не только в качестве донора электронов, но одновременно и как источник углерода. Попадая в темноту, пурпурные бактерии переходят к хемоорганогетеротрофному типу питания. При этом энергию для процессов жизнедеятельности они получают за счет реакций окисления органического субстрата по циклу Кребса. Непосредственным источником углерода и донором электронов для них являются органические соедине- ния субстрата. В клетках бактерий этой группы имеется универсальный набор дыхательных ферментов (НАД- и ФАД-дегидрогеназы и цитохромы), обеспечивающих им возможность перехода от анаэробного образа жизни на свету к аэробному в темноте и, соответственно, от авто трофного типа питания к гетеротрофному. Таким образом, среди разных групп прокариот нет, да и не может быть, резкой границы между различными типами питания.
ХемолитоавтотрофыХемолитоавтотрофы представлены микроорганизмами, способными в качестве основного источника углерода усваивать углекислый газ и синтезировать в клетке органические соединения, используя энергию реакций окисления неорганического субстрата. Для хемолитоавтотрофов неорганические вещества субстрата выступают донорами электронов в реакциях энергетического метаболизма и в процессе хемоассимиляции углекислого газа. Заслуга открытия процесса хемосинтеза принадлежит С. Н. Виноградскому. Он определил химизм как жизнь без органики. Им впервые была показана возможность образования органических веществ из неорганических в клетках бактерий, помимо процесса фотосинтеза. К хемолитоавтотрофам относится большинство видов нитрифицирующих, тионовых бактерий, некоторые виды из группы одноклеточных железобактерий и водородные бактерии. Хемолитоавтотрофные бактерии характеризуются специфичностью в отношении использования окисляемого субстрата. Нитрифицирующие бактерии для процесса хемоассимиляции углекислого газа получают энергию от окисления аммиака и нитритов. Процесс нитрификации проходит в две фазы. Первая фаза заключается в аэробном окислении аммиака до нитритов нитрозными бактериями родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira: NН3 + 11 /202 = HN02 + Н2О + 274,7 кДж. Вторая фаза нитрификации предусматривает окисление нитритов в нитраты нитратными бактериями родов Nitrobacter, Nitrospina, Nitrococcus: HN02 + 11 /202 =НNО3 + 87,5 кДж. При процессах окисления неорганического субстрата выделяется сравнительно небольшое количество энергии, и усваивается она клеткой с низким КПД, всего 5-10%. Поэтому для получения энергии на процессы жизнедеятельности нитрифицирующим бактериям приходится перерабатывать огромное количество субстрата. К хемолитоав- тотрофам относится большинство видов тионовых бактерий: Thiobacillus denitrificans, Т. thiooxidans, Т. acidophilus, Thiomicrospira pelophila и др. Они ведут процесс хемоасси- миляции углекислого газа, получая энергию за счет окисления восстановленных или частично восстановленных соединений серы, сероводорода, элементарной серы, тиосульфата и сульфита. Конечным продуктом окисления обычно является сульфат.
Некоторые виды тионовых бактерий (Thiobacillus ferrooxidans) способны получать энергию за счет окисления не только соединений серы, но и закисного железа (Fe2+) (разрушение труб). Хемолитоавтотрофный тип питания характерен для некоторых одноклеточных ацидофильных железобактерий – Leptospirillum ferrooxidans, Thiobacillus ferrooxidans и представителей рода Sulfolobus. Оптимальный рост ацидофильных железобактерий наблюдается при рН ниже 4,5 (2-3). Они используют энергию окисления Fe2+ до Fе3+ для ассимиляции С02, который служит основным или единственным источником углерода. Реакции окисления железа сопровождаются незначительным выделением энергии, поэтому железобактерии перерабатывают большие количества субстрата: 2Fe2+ + 1/202 + 2Н+ = 2FеЗ+ + Н2О + 33 кДж. Хемолитоавтотрофные бактерии в природе являются геологическими агентами. Они принимают участие в процессах образования полезных ископаемых и осуществляют важнейшие звенья круговорота азота, серы, железа. К факультативным хемолитоавтотрофам относятся водородные бактерии, являющиеся представителями 20 различных родов – Рsеudоmоnаs, Аlcаligеnеs, Nocardia и др. Они способны осуществлять ферментативное окисление водорода кислородом воздуха с образованием воды. Водородные бактерии ферментом дегидрогеназой активируют молекулярный водород и далее используют его для получения энергии и в качестве донора электронов для восстановления углекислого газа до углевода: 6Н2 + 202 + С02 = (СН2О) + 5Н2О. Являясь факультативными хемолитоавтотрофами, водородные бактерии в качестве источника энергии и углеро- да способны использовать и различные органические со- единения (сахара, органические кислоты, спирты). Таким образом, водородные бактерии могут служить еще одним 87 примером возможного перехода микробной клетки от одного типа питания к другому – от хемолитоавтотрофного к хемоорганогетеротрофному. Бактерии, переходящие от одного типа питания к другому, получили название миксо- трофов.
Хемоорганогетеротрофы. К хемоорганогетеротрофам относится большинство прокариот. Источником углерода для них являются самые разнообразные органические соединения. Энергию для жизнедеятельности они получают за счет окислительно- восстановительных реакций органического субстрата, и донором электронов в реакциях метаболизма также выступают различные органические вещества. Хемоорганогетеротрофы наиболее широко распространены в природе. Им принадлежит роль санитаров нашей планеты, так как они ведут процессы минерализации самых разнообразных, подчас сложных органических веществ. Помимо органических соединений как источника углерода, хемоорганогетеротрофы нуждаются в углекислом газе для реакций карбоксилирования промежуточного обмена. Хемоорганогетеротрофные микроорганизмы подразделяют на сапрофитов и паразитов. Сапрофиты потребляют органические вещества опада. Паразиты живут за счет органических веществ живой клетки. Выделяют факультативных и облигатных паразитов. Факультативные паразиты развиваются на обычных органических средах, но, попадая в клетку-хозяина, переходят к паразитическому образу жизни. К ним относится большинство патогенных бактерий, вызывающих заболевания человека, – возбудители пневмонии, менингита, гонореи, дизентерии, брюшного тифа, сибирской язвы, коклюша, туберкулеза и др. Облигатные (строгие) паразиты развиваются исключительно за 88 счет органических веществ клетки-хозяина. Типичным примером облигатных паразитов являются риккетсии и вирусы. В основе всех вышерассмотренных типов питания прокариот лежат различные сочетания их энергетического и конструктивного метаболизма. Всем типам питания соот- ветствуют определенные группы прокариотных организ- мов. Основной массе бактерий присущ один вполне определенный (облигатный) тип питания. Однако среди многообразного мира прокариот встречается много видов бактерий, способных переходить от одного типа питания к другому. Так, среди цианобактерий, зеленых и пурпурных бактерий имеются виды, переходящие от фотолитоавто- трофного типа питания к фотоорганогетеротрофному. Некоторые виды обширного рода Thiobacillus способны переходить от хемолитоавтотрофного типа питания к хемоорга- ногетеротрофному. Эта особенность питания прокариот обусловливает широкие возможности существования микроорганизмов в различных условиях среды и значительно расширяет их экологические ниши.
Источник



