
Типы питания прокариот. Группы микроорганизмов по типу питания
В отличие от растительных и животных организмов, имеющих один вполне определенный тип питания – соответственно автотрофный и гетеротрофный, прокариоты характеризуются многообразием типов питания. Поэтому для характеристики типов питания трокариотных организмов используются одновременно три критерия: источник углерода, источник энергии и донор электронов (водорода).
По источнику углерода прокариоты являются автотрофами, если они получают углерод в результате фиксации углекислого газа, и гетеротрофами, если источником углерода для них служат органические соединения.
По источнику энергии прокариоты, использующие солнечный свет, называются фототрофами, а получающие энергию за счет окислительно-восстановительных реакций – хемотрофами.
И наконец, по донору электронов прокариоты подразделяются на литотрофы, обладающие способностью использовать неорганические вещества, органотрофы, использующие в качестве доноров электрона органические соединения.
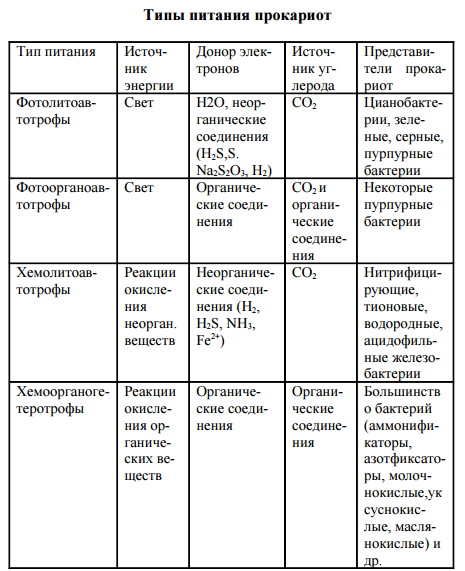
По трем вышеуказанным критериям выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы хемолитоавтотрофы и хемоорганогетеротрофы (таблица 1).
Различают следующие типы гетеротрофии: паразитизм облигатный внутриклеточный, паразитизм факультативный, сапрофитизм.
Организмы, которые могут жить только внутри других организмов, имеют редуцированный метаболизм, зависят от метаболизма хозяина, являются облигатными внутриклеточными паразитами.
Факультативные паразиты-организмы, способные расти вне клетки хозяина при подходящих условиях, на питательных средах.
Сапрофиты – гетеротрофные организмы, нуждаются в готовых органических веществах, непосредственно от организмов не зависят.
Сапрофиты нуждаются в разных концентрациях органических веществ. Олиготрофы – способны расти при низких концентрациях органического вещества (1–15 мг углерода в литре раствора). Копиотрофы – предпочитают высокие концентрации питательных веществ (10 грамм углерода в литре раствора).
Дата добавления: 2015-08-14 ; просмотров: 4897 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Типы питания прокариот
Прокариоты характеризуются многообразием типов питания в отличие от растений (автотрофы) и животных (ге- теротрофы). Для характеристики типов питания используют три критерия: источник углерода, источник энергии, донор электронов (водорода). По источнику углерода делят на ав- тотрофы и гетеротрофы, по источнику энергии – на фото- трофы (используют солнечный свет) и хемотрофы (получа- ют энергию за счет окислительно-восстановительных реак- ций), по донору – электроны (водорода) на литотрофы (неорганические соединения Н2, NH3, H2S, Fe2 + , CO и др.) и органотрофы (используют в качестве донора электрона ор- ганические соединения). Таким образом, выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы, хемолитоавтотрофы, хемоорганоге- теротрофы.
Фотолитоавтотрофы. Бактериальный фотосинтез (фоторедукция). В 1931 г. К. ван Ниль впервые доказал способность бактерий к фотосинтезу. В основе бактериального фотосинтеза лежит превращение световой энергии, поглощаемой фотосинтетическим пигментом, в биохимическую энергию макроэргических связей (АТФ) и далее использо- вание этой энергии для восстановления углекислого газа в процессе биосинтеза. У этих организмов есть пигмент бак- териохлорофилл. В клетках всех фотосинтезирующих бактерий содержатся фотосинтетические пигменты. К ним относятся особые хлорофиллы, получившие название бактериохлорофиллов а, в, с, d, и каротиноиды. По строению бактерио- хлорофиллы близки к хлорофиллу а растений. Так, бакте- риохлорофилл а отличается от хлорофилла а растений тем, что в первом его пирольном кольце в положении 2 стоит ацетильная группа СН3-СО- вместо обычной винильной группы СН2=СН-, а второе пирольное кольцо восстановле- но и содержит на два атома водорода больше. Содержание в клетке фотосинтезирующих прокариот четырех видоизменений бактериохлорофилла обусловли- вает более широкую полосу спектра поглощения световой энергии по сравнению с хлорофиллом растений. Предел спектра поглощения хлорофилла растений лежит в области 700-780 нм, в то время как фотосинтезирующие бактерии используют световую энергию длинноволновой части спектра до 1100 нм. Разница в спектрах поглощения хлоро- филлов растений и бактериохлорофиллов фотосинтезиру- ющих бактерий расширяет экологические ниши последних и позволяет фотосинтезирующим бактериям развиваться в водоемах под слоем водорослей. Помимо бактериохлорофиллов, в клетках фотосинтезирующих бактерий открыты более 20 дополнительных каро- тиноидных пигментов. Особенно разнообразны каротинои- ды в клетках пурпурных бактерий. Каротиноиды фотосин- тезирующих бактерий поглощают световую энергию ко- ротковолновой видимой области спектра с длиной волны 400-550 нм и передают эту энергию на бактериохлоро- филл. В клетке прокариот фотосинтетические пигменты нахо- дятся на инвагинациях ЦПМ – хроматофорах, визикулах (пузырьках), трубочках, тиллакоидах, ламеллах. Ван Ниль назвал этот процесс бактериальным фотосин- тезом, или фоторедукцией.
Для восстановления одной молекулы углекислого газа требуется 1 молекула АТФ (у растений 4 АТФ). В том и другом случае идет восстановление углекислого газа до углеводов. В Определителе Берджи (1974) фотосинтезирующие бактерии представлены тремя семействами (см. выше).
Фотоорганоавтотрофы Фотоорганоавтотрофы представлены немногочисленным семейством Rhodospirillaceae, включающим три рода: Rhodospirillum, Rhodopseudomonas и Rhodomicrobium. Бактерии-фотоорганоавтотрофы способны перестраивать свой обмен и одинаково успешно развиваться как на свету, так и в темноте, переходя соответственно от ана- эробного образа жизни к аэробному. На свету они ведут себя как фотоорганоавтотрофы: усваивая углекислый газ, они восстанавливают его в процессе фотосинтеза до углевода. В качестве доноров электронов несерные пурпурные бактерии используют различные органические вещества — сахара, спирты, органические кислоты, аминокислоты. Характер использования органических веществ различными фотосинтезирующими бактериями существенно раз- личается. Чаще всего органическое вещество выполняет единственную функцию донора электронов при фотоасси- миляции углекислого газа. Однако в некоторых случаях органическое вещество используется фотосинтезирующими бактериями не только в качестве донора электронов, но одновременно и как источник углерода. Попадая в темноту, пурпурные бактерии переходят к хемоорганогетеротрофному типу питания. При этом энергию для процессов жизнедеятельности они получают за счет реакций окисления органического субстрата по циклу Кребса. Непосредственным источником углерода и донором электронов для них являются органические соедине- ния субстрата. В клетках бактерий этой группы имеется универсальный набор дыхательных ферментов (НАД- и ФАД-дегидрогеназы и цитохромы), обеспечивающих им возможность перехода от анаэробного образа жизни на свету к аэробному в темноте и, соответственно, от авто трофного типа питания к гетеротрофному. Таким образом, среди разных групп прокариот нет, да и не может быть, резкой границы между различными типами питания.
ХемолитоавтотрофыХемолитоавтотрофы представлены микроорганизмами, способными в качестве основного источника углерода усваивать углекислый газ и синтезировать в клетке органические соединения, используя энергию реакций окисления неорганического субстрата. Для хемолитоавтотрофов неорганические вещества субстрата выступают донорами электронов в реакциях энергетического метаболизма и в процессе хемоассимиляции углекислого газа. Заслуга открытия процесса хемосинтеза принадлежит С. Н. Виноградскому. Он определил химизм как жизнь без органики. Им впервые была показана возможность образования органических веществ из неорганических в клетках бактерий, помимо процесса фотосинтеза. К хемолитоавтотрофам относится большинство видов нитрифицирующих, тионовых бактерий, некоторые виды из группы одноклеточных железобактерий и водородные бактерии. Хемолитоавтотрофные бактерии характеризуются специфичностью в отношении использования окисляемого субстрата. Нитрифицирующие бактерии для процесса хемоассимиляции углекислого газа получают энергию от окисления аммиака и нитритов. Процесс нитрификации проходит в две фазы. Первая фаза заключается в аэробном окислении аммиака до нитритов нитрозными бактериями родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira: NН3 + 11 /202 = HN02 + Н2О + 274,7 кДж. Вторая фаза нитрификации предусматривает окисление нитритов в нитраты нитратными бактериями родов Nitrobacter, Nitrospina, Nitrococcus: HN02 + 11 /202 =НNО3 + 87,5 кДж. При процессах окисления неорганического субстрата выделяется сравнительно небольшое количество энергии, и усваивается она клеткой с низким КПД, всего 5-10%. Поэтому для получения энергии на процессы жизнедеятельности нитрифицирующим бактериям приходится перерабатывать огромное количество субстрата. К хемолитоав- тотрофам относится большинство видов тионовых бактерий: Thiobacillus denitrificans, Т. thiooxidans, Т. acidophilus, Thiomicrospira pelophila и др. Они ведут процесс хемоасси- миляции углекислого газа, получая энергию за счет окисления восстановленных или частично восстановленных соединений серы, сероводорода, элементарной серы, тиосульфата и сульфита. Конечным продуктом окисления обычно является сульфат.
Некоторые виды тионовых бактерий (Thiobacillus ferrooxidans) способны получать энергию за счет окисления не только соединений серы, но и закисного железа (Fe2+) (разрушение труб). Хемолитоавтотрофный тип питания характерен для некоторых одноклеточных ацидофильных железобактерий – Leptospirillum ferrooxidans, Thiobacillus ferrooxidans и представителей рода Sulfolobus. Оптимальный рост ацидофильных железобактерий наблюдается при рН ниже 4,5 (2-3). Они используют энергию окисления Fe2+ до Fе3+ для ассимиляции С02, который служит основным или единственным источником углерода. Реакции окисления железа сопровождаются незначительным выделением энергии, поэтому железобактерии перерабатывают большие количества субстрата: 2Fe2+ + 1/202 + 2Н+ = 2FеЗ+ + Н2О + 33 кДж. Хемолитоавтотрофные бактерии в природе являются геологическими агентами. Они принимают участие в процессах образования полезных ископаемых и осуществляют важнейшие звенья круговорота азота, серы, железа. К факультативным хемолитоавтотрофам относятся водородные бактерии, являющиеся представителями 20 различных родов – Рsеudоmоnаs, Аlcаligеnеs, Nocardia и др. Они способны осуществлять ферментативное окисление водорода кислородом воздуха с образованием воды. Водородные бактерии ферментом дегидрогеназой активируют молекулярный водород и далее используют его для получения энергии и в качестве донора электронов для восстановления углекислого газа до углевода: 6Н2 + 202 + С02 = (СН2О) + 5Н2О. Являясь факультативными хемолитоавтотрофами, водородные бактерии в качестве источника энергии и углеро- да способны использовать и различные органические со- единения (сахара, органические кислоты, спирты). Таким образом, водородные бактерии могут служить еще одним 87 примером возможного перехода микробной клетки от одного типа питания к другому – от хемолитоавтотрофного к хемоорганогетеротрофному. Бактерии, переходящие от одного типа питания к другому, получили название миксо- трофов.
Хемоорганогетеротрофы. К хемоорганогетеротрофам относится большинство прокариот. Источником углерода для них являются самые разнообразные органические соединения. Энергию для жизнедеятельности они получают за счет окислительно- восстановительных реакций органического субстрата, и донором электронов в реакциях метаболизма также выступают различные органические вещества. Хемоорганогетеротрофы наиболее широко распространены в природе. Им принадлежит роль санитаров нашей планеты, так как они ведут процессы минерализации самых разнообразных, подчас сложных органических веществ. Помимо органических соединений как источника углерода, хемоорганогетеротрофы нуждаются в углекислом газе для реакций карбоксилирования промежуточного обмена. Хемоорганогетеротрофные микроорганизмы подразделяют на сапрофитов и паразитов. Сапрофиты потребляют органические вещества опада. Паразиты живут за счет органических веществ живой клетки. Выделяют факультативных и облигатных паразитов. Факультативные паразиты развиваются на обычных органических средах, но, попадая в клетку-хозяина, переходят к паразитическому образу жизни. К ним относится большинство патогенных бактерий, вызывающих заболевания человека, – возбудители пневмонии, менингита, гонореи, дизентерии, брюшного тифа, сибирской язвы, коклюша, туберкулеза и др. Облигатные (строгие) паразиты развиваются исключительно за 88 счет органических веществ клетки-хозяина. Типичным примером облигатных паразитов являются риккетсии и вирусы. В основе всех вышерассмотренных типов питания прокариот лежат различные сочетания их энергетического и конструктивного метаболизма. Всем типам питания соот- ветствуют определенные группы прокариотных организ- мов. Основной массе бактерий присущ один вполне определенный (облигатный) тип питания. Однако среди многообразного мира прокариот встречается много видов бактерий, способных переходить от одного типа питания к другому. Так, среди цианобактерий, зеленых и пурпурных бактерий имеются виды, переходящие от фотолитоавто- трофного типа питания к фотоорганогетеротрофному. Некоторые виды обширного рода Thiobacillus способны переходить от хемолитоавтотрофного типа питания к хемоорга- ногетеротрофному. Эта особенность питания прокариот обусловливает широкие возможности существования микроорганизмов в различных условиях среды и значительно расширяет их экологические ниши.
Источник
Питание прокариот. Химический состав клетки прокариот
» data-shape=»round» data-use-links data-color-scheme=»normal» data-direction=»horizontal» data-services=»messenger,vkontakte,facebook,odnoklassniki,telegram,twitter,viber,whatsapp,moimir,lj,blogger»>
ПИТАНИЕ ПРОКАРИОТ
ХИМИЧЕСКИЙ СОСТАВ ПРОКАРИОТНОЙ КЛЕТКИ
Вода в клетке прокариот составляет 80—90% от обшей массы.
На долю сухого вещества в прокариотной клетке приходится 10—20% от общей массы.
Элементарный состав сухих веществ прокариотной клетки характеризуется следующими данными (в % к сухой массе):
углерод— 50, кислород — 20, азот— 10—— 15, водород— 10, фосфор — 2—6, остальную часть составляют сера и прочие элементы (К, Na, Ca, Mg, Fe, Mn, Mo и др.).
На долю белков приходится 50—80% сухой массы, углеводы -3-10%, липиды – 3-4%.
По химическому составу белки прокариот почти не отличаются от белков эукариот. В состав белков прокариот входят 20 аминокислот. Из специфических аминокислот в клетках бактерий обнаружены диаминопимелиновая и диаминомасляная кислоты.
Нуклеиновые кислоты в клетке прокариот представлены ДНК и РНК- Последняя сосредоточена в цитоплазме и составляет около 16% сухой массы. ДНК образует нуклеоид и в клетках некоторых бактерий плазмиды. На долю ДНК приходится примерно 3—4% cyxoй массы.
Углеродное питание Бактерий.
По источнику углерода для конструктивного обмена все прокариоты делятся на две группы: автотрофы, потребляющие в качестве главного источника углерода углекислый газ, и гетеротрофы, усваивающие углерод из органических соединений.
Автотрофы делятся на фототрофов, фоторедукторов, хемотрофов.
1. Фототрофы.
Синтезируют органическое вещество из СО2 и Н2О с помощью энергии солнца. Фотосинтез идет обычным путем. Процесс фотосинтеза осуществляется 2 группами бактерий: цианобактериями и прохлорофитами. У них есть н6абор пигментов.
Для большинства гетеротрофов оптимальным и наиболее доступным источником углерода служат углеводы. Рассмотрим хемосинтез на примере азотофиксирующих бактерий. Они обитают в почве и окисляют NH3 до HNO3, а затем до HNO2.
Гетеротрофы делятся на сапрофитов и паразитов. Сапрофиты используют С мертвых органических остатков. Паразиты – питаются орг. Соед. Живых организмов. Паразиты бывают облигатные (обязательные – вирусы, риккетсии) и факультативные (все виды болезнетворных бактерирй).
Помимо углеводов, хорошим источником углерода для многих бактерий являются многоатомные спирты и аминокислоты. Некоторые виды прокариот способны усваивать углерод из органических кислот.
Азот
Для синтеза аминокислот, пуриновых и лйримидииовых нуклеотидов бактериям необходим азот. В природе азот встречается в форме окисленных и восстановленных соединений, а также в виде молекулярного азота атмосферы.
Большинство прокариот потребляют азот в восстановленной форме в виде солей аммония (NH4+) и аммиака (NH3). Многие бактерии используют органические азотсодержащие вещества — белки, аминокислоты, мочевину, разрушая их с выделением аммиака. Окисленные формы азота — нитриты, нитраты — также усваиваются различными группами бактерий. Среди прокариот известно большое число организмов — бактерий, актиномицетов, сине-зеленых водорослей, способных фиксировать молекулярный азот атмосферы для построения всех необходимых компонентов клетки.
Фосфор в клетках прокариот входит в состав важнейших органических соединений — нуклеиновых кислот, фосфолипидов, ко-ферментов. Такие соединения фосфора, как АДФ и АТФ, являются аккумуляторами энергии клетки и играют важную роль в метаболизме. Источником фосфора для бактерий в основном служат фосфаты калия или натрия, а из органических соединений нуклеиновые кислоты.
Сера в клетке прокариот в основном встречается в восстановленной форме и входит в состав аминокислот, витаминов и кофакторов (биотин, кофермент А и др.). Наиболее важным компонентом, содержащим серу, является цистеин. Атомы серы в большинстве других содержащих серу соединений клетки (метио-нин, биотин, тиамин) происходят из SH-группы цистеина. Источником серы для большинства микроорганизмов служат сульфаты, которые в клетке восстанавливаются в сульфиды. Некоторые бактерии нуждаются в соединениях, содержащих серу в восстановленной форме, таких, как сероводород, тиосульфат, цистеин и метионин.
Для нормального роста и развития прокариот необходимы ионы металлов, представленные макроэлементами, такими, как калий, кальций, магний, железо, и микроэлементами. К последним относятся марганец, молибден, цинк, медь, кобальт, никель и др. Ионы металлов входят в состав жизненно важных метаболитов бактериальной клетки. Так, кобальт является активатором ферментов транспорта электронов в окислительно-восстановительных реакциях цикла Кребса. Железо и молибден необходимы бактериям для синтеза ферментов, участвующих в процессе азот-фиксации.
Особого внимания заслуживает магний, так как, помимо активации ферментов, таких, как гексокиназа, он определяет агрегацию мономеров рибосомы.
Факторы роста, факторами роста называются органические соединения, которые не синтезируются многими прокариотными организмами, но без которых жизнь клетки оказывается невозможна. К таким соединениям относятся аминокислоты, пурины, пиримидины, витамины и др. Эти соединения прокариоты должны получать из среды.
Бактерии, нуждающиеся в каком-либо факторе роста, называются ауксотрофными по отношению к этому соединению, в отличие от прототрофных, способных синтезировать данное вещество в клетке.
Прокариоты существенно различаются по потребностям в факторах роста. Например, молочнокислые бактерии ауксотрофны ко многим аминокислотам, пуринам, пиримидинам и 5—б витаминам, в то время как различные штаммы Escherichia coll проявляют ауксотрофность к какому-либо одному, но разному фактору роста.
МЕХАНИЗМ ПОСТУПЛЕНИЯ ПИТАТЕЛЬНЫХ ВЕЩЕСТВ В КЛЕТКУ ПРОКАРИОТ
Избирательное поступление веществ питательного субстрата в клетку прокариот регулируется цитоплазматической мембраной. ь Клеточная стенка служит вторым барьером на пути проникновения веществ в клетку. Она задерживает некоторые крупномолекулярные соединения питательного субстрата, например такие, как декстраны. У некоторых бактерий клеточная стенка обладает избирательной проницаемостью для низкомолекулярных соединений, основанной на взаимном притяжении разнозаряженных частиц.
Цитоплазматическая мембрана клетки несет ответственность за поступление воды и веществ питательного субстрата в клетку и определяет выход продуктов обмена наружу. В настоящее время изучено несколько механизмов переноса веществ субстрата через цитоплазматическую мембрану в клетку прокариот.
Пассивная диффузия — процесс поступления воды и некоторых растворенных веществ через цитоплазматическую мембрану в цитоплазму клетки по градиенту концентрации, от большей концентрации к меньшей для неэлектролитов или по градиенту электрических потенциалов для ионов. Скорость пассивной диффузии невелика, и проходит она без затраты энергии.L
Облегченная диффузия отличается от пассивной тем, что перенос веществ субтрата через цитоплазматическую мембрану осуществляется белками-переносчиками, получившими название пермеаз (транслоказ). Пермеаза катализирует присоединение вещества субстрата к активному центру на своей поверхности и проводит это вещество с наружной поверхности цитоплазматической мембраны на внутреннюю. Там пермеаза освобождается от этого вещества, передавая его в цитоплазму, а сама вновь вступает во взаимодействие с новой порцией субстрата.
Пермеазные белки синтезируются и локализуются на цитоплазматической мембране.
Активный транспорт заключается в переносе веществ субстрата пермеазньми белками через цитоплазматическую мембрану в цитоплазму против градиента концентрации. Процесс активного транспорта веществ всегда осуществляется с затратой энергии. Учитывая, что на перенос одной молекулы субстрата через цитоплазматическую мембрану клетка расходует одну молекулу АТФ, можно предполагать, что растущий и быстро размножающийся микроорганизм затрачивает значительную часть вырабатываемой энергии на транспорт веществ.
Необходимым условием поступления веществ субстрата в клетку бактерий является их растворимость в воде.
Вещества питательного субстрата, поступающие в клетку, являются источником энергетического метаболизма и одновременно строительным материалом для синтеза клеточных структур. Выход продуктов обмена из клетки осуществляется чаще всего путем облегченной диффузии при участии белков-переносчиков.
ТИПЫ ПИТАНИЯ ПРОКАРИОТ
В отличие от растительных и животных организмов, имеющих один вполне определенный тип питания — соответственно автотрофный и гетеротрофный, прокариоты характеризуются многообразием типов питания. Поэтому для характеристики типов питания прокариотных организмов используются одновременно три критерия; источник углерода, источник энергии и донор электронов (водорода).
Как указывалось выше, по источнику углерода прокариоты являются автотрофами, если они получают углерод в результате фиксации углекислого газа, и гетеротрофами, если источником углерода для них служат органические соединения.
По источнику энергии прокариоты, использующие солнечный свет, называются фототрофами, а получающие энергию за счет окислительно-восстановительных реакций — хемотрофами.
И наконец, по донору электронов прокариоты подразделяются на литотрофы, обладающие способностью использовать неорганические доноры электронов (H2S, Fe 2 +, СО и т. д.), и органотрофы, использующие в качестве доноров электронов органические соединения.
По трем вышеуказанным критериям выделяют 4 основных типа питания прокариот: фотолитоавтотрофы, фотоорганоавтотрофы, хемолитоавтотрофы и хемоорганогетеротрофы (табл. 6).
| Тип питания | Источник углерода | Источник | Донор электронов | Представители прокариот |
| Фотолитоавтотрофы | СО2 | Свет | Н2О Неорганические соединении (H2S, S, Na2S) | Цианобактерии. Зеленые, серные пурпурные бактерии |
| Фотоорганоавтотрофы | СО2 и органические соединения | Свет | Органические соединения (спирты, органические кислоты и др.) | Некоторые пурпурные бактерии |
| Хемолитоавтотрофы | СО, | Реакции окисления неорганических веществ | Неорганические соединении (Н2, H,S, NH2, Fe2+ и др.) | Нитрифицирующие, тионовые, водородные бактерии; ацидофильные железобактерии |
| Хемоорганогетеротрофы | Органические соединения | Реакции окисления opганических веществ | Органические соединения | Большинство бактерий (аммонификаторы, азотфиксаторы, пектино-разрушающие, клетчат-коразрушающие, молочнокислые, уксуснокислые, маслянокислые и ДР-) |
Фотолитоавтотрофы. Бактериальный фотосинтез.
К группе фотолитоавтотрофов относятся прокариоты, использующие в качестве источника углерода CO2, а в качестве донора электронов различные неорганические соединения (Н2О, H2S, S и др.). Усваивая энергию солнечного света в процессе фотосинтеза, они образуют органические вещества клетки (табл. 6). Фотолитоавтотрофы представлены цианобактериями (сине-зелеными водорослями), зелеными и серными-пурпурными бактериями.
В основе процесса бактериального фотосинтеза лежит превращение световой энергии, поглощаемой фотосинтетическими пигментами, в биохимическую энергию макроэргических связей (АТФ) и далее использование этой энергии для усвоения и восстановления углекислого газа в процессе биосинтеза.
В клетках всех фотосинтезирующих бактерий содержатся фотосинтетические пигменты. К ним относятся особые хлорофиллы, получившие название бактериохлорофиллов а, b, с, d, и каротиноиды. По строению бактериохлорофиллы близки к хлорофиллу а растений.
Помимо бактериохлорофиллов в клетках фотосинтезирующих бактерий открыты более 20 дополнительных каротиноидных пигментов.
В клетке прокариот фотосинтегические пигменты локализуются на внутриклеточных инвагинациях цитоплазматической мембраны, получивших название хроматофоров. У разных типов бактерий инвагинации иитоплазматической мембраны имеют различную форму — пузырьков (везикул), трубочек и тилакоидов, образованных стопками ламелл, напоминающих тилакоиды хлоропластов растений (рис. 2).
По химизму фотосинтез бактерий существенно отличается от фотосинтеза растений. При фотосинтезе растений и цианобактерий донором электронов является вода и процесс фотосинтеза обязательно сопровождается выделением кислорода. Причем растительная клетка для восстановления одной молекулы углекислого газа потребляет 4 кванта энергии:
CO2 + H2O — (CH2O) + O2
В процессе фотосинтеза зеленых и пурпурных бактерий в качестве доноров электронов выступают различные соединения: сероводород, элементарная сера, сульфит, тиосульфат, молекулярный водород и органические вещества. Кислород при фотосинтезе зеленых и пурпурных бактерий не выделяется. Они являются облигатными анаэробами, исключение составляет лишь немногочисленная группа несерных пурпурных бактерий, относящихся к факультативным анаэробам. Для восстановления одной молекулы углекислого газа зеленые и пурпурные бактерии затрачивают один квант энергии:
где Н2А — донор водорода.
Суть процессов бактериального фотосинтеза и фотосинтеза растений принципиально одинакова и заключается в восстановлении углекислого газа до соединения типа углеводов (СНэО).
Представители семейств Chlorobacteriaceae и Chrornatiaceae по типу питания преимущественно являются фотолитоавтотрофами. В качестве донора электронов они чаще всего используют сероводород, окисляя его до свободной серы или до сульфатов.
У зеленых бактерий сера обычно выделяется в среду, у серных пурпурных бактерий — откладывается в клетке. При отсутствии сероводорода в среде серные пурпурные и зеленые бактерии способны окислять элементарную серу, используя ее источником электронов.
Помимо сероводорода и серы зеленые и серные пурпурные бактерии донором электронов используют такие соединения, как сульфит H2SO3, тиосульфат Na
Фотоорганоавтотрофы.
Фотоорганоавтотрофы представлены немногочисленным семейством Rhodospirillaceae, включающим три рода: Rhodospirillum, Rhodopscudomonas и Rhodomicrobium.
Бактерии-фотоорганоавтотрофы способны перестраивать свой обмен и одинаково успешно развиваться как на свету, так и в темноте, переходя соответственно от анаэробного образа жизни к аэробному. На свету они ведут себя как фотоорганоавтотрофы: усваивая углекислый газ, они восстанавливают его в процессе фотосинтеза до углевода, В качестве доноров электронов несерные пурпурные бактерии используют различные органические вещества — сахара, спирты, органические кислоты, аминокислоты (табл. 6). Например, бактерии Rhodopseudomonas используют в процессе фотосинтеза донором электронов изопропанол, окисляя его в ацетон.
Характер использования органических веществ различными фотосинтезирующими бактериями существенно различается. Чаще всего органическое вещество выполняет единственную функцию — донора электронов при фотоассимиляции углекислого газа. Однако в некоторых случаях органическое вещество используется фотосинтезирующими бактериями не только в качестве донора электронов, но одновременно и как источник углерода.

Рис. 23. Фотосинтезирующие бактерии. I — Rhodospirillum; 2—Rhodomicrobium; 3 — Chromatium; 4 — Tliiopedia; 5 —Chlorobium, 6— Pelodictyon.
Попадая в темноту, несерные пурпурные бактерии переходят к хемоорганогетеротрофному типу питания. При этом энергию для процессов жизнедеятельности они получают за счет реакций окисления органического субстрата по циклу Кребса. Непосредственным источником углерода и донором электронов для них являются органические соединения субстрата.
Хемолитоавтотрофы.
Хемолитоавтотрофы представлены микроорганизмами, способными в качестве основного источника углерода усваивать углекислый газ и синтезировать в клетке органические соединения, используя энергию реакций окисления неорганического субстрата. Для хемолитоавтотрофов неорганические вещества субстрата выступают донорами электронов в реакциях энергетического метаболизма и в процессе хемоассимиляции углекислого газа (табл. 6).
Заслуга открытия процесса хемосинтеза принадлежит С. Н. Виноградскому.
К хемолитоавтотрофам относится большинство видов нитрифицирующих, тионовых бактерий, некоторые виды из группы одноклеточных железобактерий и водородные бактерии.
Хемолитоавтотрофные бактерии характеризуются специфичностью в отношении использования окисляемого субстрата. Нитрифицирующие бактерии для процесса хемоассимиляции углекислого газа получают энергию от окисления аммиака и нитритов. Процесс нитрификации проходит в две фазы. Первая фаза заключается в аэробном окислении аммиака до нитритов нитрозными бактериями родов Nitrosomonas, Nitrosococcus, Nitrosolobus, Nitrosospira.
Вторая фаза нитрификации предусматривает окисление нитритов в нитраты нитратными бактериями родов Nitrobacter, Nitrospina, Nitrococcus.
При процессах окисления неорганического субстрата выделяется сравнительно небольшое количество энергии, и усваивается она клеткой с низким КПД, всего 5—10%. Поэтому для получения энергии на процессы жизнедеятельности нитрифицирующим бактериям приходится перерабатывать огромное количество субстрата.
К хемолитоавтотрофам относится большинство видов тионовых бактерий:. Они ведут процесс хемоассимиляции углекислого газа, получая энергию за счет окисления восстановленных или частично восстановленных соединений серы, сероводорода, элементарной серы, тиосульфата и сульфита. Конечным продуктом окисления обычно является сульфат.
Хемолитоавтотрофный тип питания характерен для некоторых одноклеточных ацидофильных железобактерий —Они используют энергию окисления Fe 2+ до Fe 4+ для ассимиляции СО2, который служит основным или единственным источником углерода. Реакции окисления железа сопровождаются незначительным выделением энергии, поэтому железобактерии перерабатывают большие количества субстрата.
Хемолитоавтотрофные бактерии в природе являются геологическими агентами. Они принимают участие б процессах образования полезных ископаемых и осуществляют важнейшие звенья круговорота азота, серы, железа.
Хемоорганогетеротрофы.
К хемоорганогстеротрофам относится большинство прокариот. Источником углерода для них являются самые разнообразные органические соединения. Энергию для жизнедеятельности они получают за счет окислительно-восстановительных реакций органического субстрата, и донором электронов в реакциях метаболизма также выступают различные органические вещества (табл. 6).
Хемоорганогетеротрофы наиболее широко распространены в природе. Им принадлежит роль санитаров нашей планеты, так как они ведут процессы минерализации самых разнообразных, подчас сложных органических веществ. Помимо органических соединений, как источника углерода, хемоорганогетеротрофы нуждаются в углекислом газе для реакций карбоксилирования промежуточного обмена.
Хемоорганогетеротрофные микроорганизмы подразделяют на сапрофитов и паразитов. Сапрофиты потребляют органические вещества опада. Паразиты живут за счет органических веществ живой клетки. Выделяют факультативных и облигатных паразитов. Факультативные паразиты развиваются на обычных органических средах, но, попадая в клетку-хозяина, переходят к паразитическому образу жизни. К ним относится большинство патогенных бактерий, вызывающих заболевания человека,— возбудители пневмонии, менингита, гонореи, дизентерии, брюшного тифа, сибирской язвы, коклюша, туберкулеза и др. Облигатные (строгие) паразиты развиваются исключительно за счет органических веществ клетки-хозяина. Типичным примером облигатных паразитов являются риккетсии и вирусы.
Источник



