
Способы передвижения у птиц

Птицы приспособились передвигаться по самым различным субстратам. Они хорошо передвигаются по земле, лазают по деревьям и отвесным скалам, плавают и ныряют. Но самым характерным способом передвижения является полет. Нелетающих птиц среди современных видов немного. К ним принадлежат страусоподобные птицы, киви, а также некоторые островные виды, утратившие способность к полету вторично.
Характер полета отдельных групп птиц весьма разнообразен и находится в зависимости как от экологических условий обитания птиц, так и от их строения. Организация крыла, отношение веса тела к площади крыльев, развитие мускулатуры – вот основные факторы, определяющие свойства полета у птиц. Выделяют два основных способа полета у птиц – машущий и парящий.
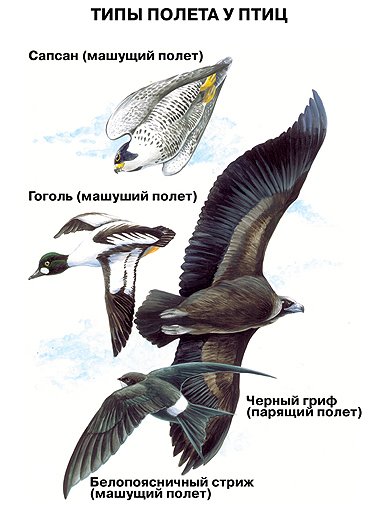
Машущий полет еще называют активным, а парящий полет – пассивным. Во время машущего полета птица совершает движения крыльями. Он сложнее и разнообразнее парящего. Движения крыльев во время машущего полета могут различаться как по частоте, так и по силе. Стоит только сравнить стремительный полет стрижа, медлительный полет вороны, трепещущую в воздухе пустельгу, быстро летящую утку или тяжело хлопающего крыльями фазана, чтобы понять все многообразие способов активного полета. Предпринимались неоднократные попытки его классификации, но общепризнанной системы выработано так и не было. К машущему полету относится и пикирование, при помощи которого многие птицы настигают жертву. Парящий полет принципиально отличается от машущего тем, что птица использует восходящие потоки воздуха, которые образуются вследствие неравномерного нагрева поверхности земли солнцем. Во время парящего полета птица практически не совершает движения крыльями. Используя разности температуры различных потоков воздуха, неравномерное действие ветра и другие факторы, птица может часами парить, практически не прикладывая никаких усилий. Для парителей характерны крупные размеры, длинные крылья с развитой несущей поверхностью, относительно малые размеры сердца. Крыло может быть либо широким (наземные виды), либо узким (морские виды).

По земле птицы передвигаются двумя основными способами. Те виды, которые большую часть времени проводят на земле, относятся к экологической группе наземных птиц. Для них характерно передвижение по субстрату шагом или бегом. Шагом передвигаются по земле и птицы, предпочитающие водный образ жизни. Но наличие перепонок на лапах делает их походку неуклюжей. Гагары же вообще с трудом передвигаются по суше, часто просто переползают, опираясь на брюхо и отталкиваясь задними конечностями. Поэтому гнездо гагар располагается прямо у уреза воды, чтобы минимизировать время, необходимое на перемещение по суше. Древесные виды, конечности которых приспособлены в большей степени к перемещению по ветвям деревьев, передвигаются по земле прыжками. Существуют птицы, которые вообще не могут передвигаться по поверхности земли. Ноги их настолько короткие, что с их помощью можно только цепляться за кору деревьев или каменистые склоны. Такие виды формируют экологическую группу воздухореев, к которой относятся стрижи.
Большое число видов птиц приспособились к водному образу жизни. Многие пернатые превосходно плавают и ныряют. Для движения в воде используются задние конечности. При этом происходят преобразования, направленные на увеличение поверхности стопы, чтобы увеличить силу отталкивания от воды. У разных видов они проходят по-разному.

У трубконосых, гагар, гусеообразных, чаек и чистиков три передних пальца соединены кожистой перепонкой. У веслоногих такая же перепонка соединяет все четыре пальца. У лысухи по краям пальцев формируются фигурные фестоны, у поганок пальцы уплощаются и формируют плавательные лопасти. У некоторых куликов (плавунчики) формируются небольшие перепонки также между передними тремя пальцами. Некоторые водные и околоводные птицы могут превосходно нырять. При этом под водой в качестве основного движителя используются либо задние, либо передние конечности. Чистики под водой совершают крыльями движения, похожие на полет. Напротив, нырковые утки, гагары, поганки, лысухи передвигаются под водой с помощью задних конечностей. У уток крылья могут использоваться только как своеобразные рули глубины. Для некоторых видов характерно ныряние с большой высоты. Они не плавают под водой, а погружаются на краткие мгновения лишь для того, чтобы схватить добычу. К таким птицам в наших широтах относятся скопы, зимородки, крачки и некоторые другие виды.
Источник
Двигательная система птиц и основные типы движения
Движения птиц многообразны: ходьба, прыжки, бег, лазанье, плавание, ныряние, полет. Они обеспечиваются как изменениями опорно-мышечной системы, так и преобразованиями других систем органов, осуществляющих координацию движений и ориентировку в пространстве, создающих необходимые энергетические резервы. Своеобразная особенность скелета птиц — хорошо выраженная пневматичность костей. Плоские кости имеют губчатое строение, сохраняя большую прочность при небольшой толщине. Трубчатые кости тоже тонкостенны, а полости внутри них заполнены частично воздухом, частично — костным мозгом. Эти особенности обеспечивают повышенную прочность отдельных костей и заметно их облегчают.
Нужно, однако, обратить внимание на то, что общая масса скелета составляет 8-18% массы тела птиц — примерно столько же, сколько у млекопитающих, у которых кости толще, а воздушные полости в них отсутствуют. Это объясняется тем, что у птиц облегчение костей позволило резко увеличить их длину (длина скелета ноги, а особенно крыла в несколько раз превышает длину туловища), заметно не повышая общей массы скелета.
Как и у других высших позвоночных, скелет птиц подразделяется на осевой- скелет и связанную с ним грудную клетку, череп, скелет конечностей и их поясов.
Осевой скелет-позвоночный столб подразделяется на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой. Число шейных позвонков изменчиво — от 11 до 23-25 (лебеди). Как и у пресмыкающихся, первый позвонок — атлас, или атлант, — имеет форму костного кольца, а второй — эпистрофей — сочленяется с ним зубовидным отростком; это обеспечивает подвижность головы относительно шеи. Остальные шейные позвонки птиц гетероцельного типа, длинное тело каждого позвонка спереди и сзади имеет седлообразную поверхность (в сагиттальном разрезе позвонки опистоцельны, а во фронтальном — предельны). Сочленение таких позвонков обеспечивает их значительную подвижность относительно друг друга в горизонтальной и вертикальной плоскостях. Прочность соединений позвонков усиливается наличием суставных отростков на основаниях верхних дуг, образующих между собой скользящие суставы.
Шейные ребра птиц рудиментарны и срастаются с шейными позвонками, образуя канал, по которому проходят позвоночная артерия и шейный симпатический нерв. Только последние одно-два шейных ребра сочленяются с шейными позвонками подвижно, однако до грудины они не доходят. Особенности шейных позвонков вместе со сложно дифференцированными шейными мышцами позволяют птицам свободно поворачивать голову на 180°, а некоторым «(совы, попугаи) и на 270°. Это делает возможным сложные и быстрые движения головой при схватывании подвижной добычи, чистке оперения, постройке гнезда; в полете позволяет, сгибая или разгибая шею, в определенных пределах менять положение центра тяжести, облегчает ориентировку и т. п.
Грудных позвонков у птиц 3-10. Они срастаются друг с другом, образуя спинную кость, и очень тугим суставом соединяются со сложным крестцом. Благодаря этому туловищный отдел осевого скелета становится неподвижным, что важно при полете (колебания туловища не мешают координации летательных движений). К грудным позвонкам подвижно причленяются ребра. Каждое ребро состоит из двух отделов — спинного и брюшного, подвижно сочленяющихся друг с другом и образующих угол, вершиной направленный назад. Верхний конец спинного отдела ребра подвижно причленяется к поперечному отростку и телу грудного позвонка, а нижний конец брюшного отдела — к краю грудины. Подвижное сочленение спинного и брюшного отделов ребер между собой и подвижное их соединение с позвоночным столбом и грудиной наряду с развитой реберной мускулатурой обеспечивают изменение объема полости тела. Это один из механизмов интенсификации дыхания. Прочность грудной клетки усиливается крючковидными отростками, укрепленными на спинных отделах и налегающих на последующее ребро. Большая грудина имеет вид тонкой широкой и длинной пластинки, на которой у всех птиц (кроме страусоподобных) расположен высокий киль грудины. Большие размеры грудины и ее киля обеспечивают место для прикрепления мощных мышц, двигающих крыло.
Все поясничные, крестцовые (их два) и часть хвостовых позвонков неподвижно срастаются друг с другом в монолитную кость — сложный крестец. Всего в него входят 10-22 позвонка, границы между которыми не видны. Со сложным крестцом неподвижно срастаются кости тазового пояса. Это обеспечивает неподвижность туловищного отдела и создает прочную опору для задних конечностей. Число свободных хвостовых позвонков не превышает 5-9. Последние 4-8 хвостовых позвонков сливаются в уплощенную с боков копчиковую кость, к которой веером прикрепляются основания рулевых перьев. Укорочение хвостового отдела и образование пигостиля обеспечивает прочную опору хвосту при сохранении его подвижности. Это важно, так как хвост не только выполняет функцию добавочной несущей плоскости, но участвует и в управлении полетом (как тормоз и руль).
Череп птиц похож на череп рептилий и может быть отнесен к диапсидному типу с редуцированной верхней дугой. Череп тропибазальный (глазницы расположены впереди головного мозга), образован тонкими губчатыми костями, границы между которыми отчетливо видны лишь у молодых птиц. Это, видимо, связано с тем, что соединение при помощи швов невозможно из-за небольшой толщины костей. Поэтому череп относительно легок. Своеобразна по сравнению с пресмыкающимися и его форма: резко увеличен объем мозговой коробки, глазницы большие, челюсти лишены зубов (у современных птиц) и формируют клюв. Смещение большого затылочного отверстия и затылочного мыщелка на дно черепа увеличивает подвижность головы относительно шеи и туловища.
Большое затылочное отверстие окружено четырьмя затылочными костями: основной, двумя боковыми и верхней. Основная и боковые затылочные кости образуют единственный (как и у пресмыкающихся) затылочный мыщелок, сочленяющийся с первым шейным позвонком. Три ушные кости, окружающие слуховую капсулу, сливаются с прилегающими костями и между собой. В полости среднего уха находится лишь одна слуховая косточка — стремечко. Бока и крышу мозговой коробки образуют парные покровные кости: чешуйчатые, теменные, лобные и боковые клиновидные. Дно черепа образует покровная основная клиновидная кость, которую закрывает покровная основная височная кость, и клювовидный отросток парасфеноида. У его переднего конца лежит сошник, по краям которого расположены хоаны.
Верхняя часть клюва — надклювье — образована сильно разросшимися и слившимися предчелюстными костями. Гребень клюва, укрепленный носовыми костями, соединяется с лобными костями и передней стенкой глазницы, образованной разросшейся средней обонятельной костью. Составляющие лишь заднюю часть надклювья верхнечелюстные кости отростками сливаются с небными костями. К задненаружному краю верхнечелюстной кости прирастает тонкая костная перекладина, состоящая из двух слившихся костей — скуловой и квадратно-скуловой. Это типичная нижняя дуга диапсидного черепа, ограничивающая снизу глазницу и височную яму. Квадратно-скуловая кость сочленяется с квадратной костью, нижний конец которой образует суставную поверхность для сочленения с нижней челюстью, а удлиненный верхний конец суставом прикрепляется к чешуйчатой и переднеушной костям. Небные кости концами налегают на клювовидный отросток парасфеноида и суставом соединяются с парными крыловидными костями, которые в свою очередь суставом связаны с квадратными костями соответствующей стороны.

Такое строение костного неба имеет важное значение для свойственного большинству птиц кинетизма (подвижности) надклювья. При сокращении мышц, соединяющих направленный вперед глазничный отросток квадратной кости со стенкой глазницы, нижний конец квадратной кости смещается вперед и сдвигает как небные и крыловидные кости (их соединение друг с другом может скользить по клювовидному отростку), так и квадратно-скуловые и скуловые. Давление по этим костным мостикам передается на основание надклювья и благодаря, перегибу костей в области «переносицы» вершина надклювья сдвигается кверху. В зоне перегиба надклювья кости очень тонки, а у некоторых видов (гуси и др.) здесь образуется сустав. При сокращении мышц, соединяющих череп с нижней челюстью, вершина клюва сдвигается книзу. Подвижность костного неба в сочетании со сложно дифференцированными жевательными мышцами обеспечивают разнообразные, тонко дифференцированные движения клюва при схватывании добычи, чистке оперения, постройке гнезд. Вероятно, подвижность шеи и приспособление клюва к многообразным движениям способствовали превращению передних конечностей в крылья, так как замещали некоторые из выполняемых ими второстепенных функций (помощь в захвате пищи, чистка тела и др.).
Нижняя часть клюва — подклювье или нижняя челюсть — образуется слиянием ряда костей, из которых более крупные зубная, сочленовная и угловая. Челюстной сустав формируют сочленовная и квадратная кости. Движения надклювья и подклювья очень четко координированы благодаря дифференцированной системе жевательных мышц. Подъязычный аппарат состоит из удлиненного тела, поддерживающего основание языка, и длинных рожек. У некоторых птиц, например дятлов, очень длинные рожки огибают весь череп. При сокращении подъязычной мускулатуры рожки скользят по соединительнотканному ложу и язык выдвигается из ротовой полости почти на длину клюва.
Скелет передней конечности, превратившейся у птиц в крыло, подвергся значительным изменениям. Мощная трубчатая кость — плечо — имеет уплощенную головку, что существенно ограничивает вращательные движения в плечевом суставе, обеспечивая устойчивость крыла в полете. Дистальный конец плеча сочленяется с двумя костями предплечья: более прямой и тонкой лучевой и более мощной локтевой, на задневерхней стороне которой видны бугорки — места прикрепления очинов второстепенных маховых. Из проксимальных элементов запястья сохраняются лишь две маленькие самостоятельные косточки, которые связками соединяются с костями предплечья. Кости дистального ряда запястья и все кости пясти сливаются в общую пястно-запястную кость, или пряжку. Резко редуцируется скелет пальцев: хорошо развиты только две фаланги второго пальца, продолжающие ось пряжки. От первого и третьего пальца сохраняется лишь по одной короткой фаланге. Первостепенные маховые прикрепляются к пряжке и к фалангам второго пальца. К фаланге первого пальца прикрепляется несколько перьев «крылышка».
Преобразование кисти (образование пряжки, редукция пальцев, малая подвижность сустава) обеспечивают прочную опору первостепенным маховым, испытывающим в полете наибольшие нагрузки. Характер поверхностей всех суставов таков, что обеспечивает легкую подвижность лишь в плоскости крыла; возможность вращательных движений резко ограничена. Это предохраняет выворачивание крыла, позволяет птице без усилий менять площадь крыла в полете и складывать его в покое. Соединяющая кистевой сгиб с плечевым суставом складка кожи — летательная перепонка — образует эластичный передний край крыла, сглаживающий локтевой сгиб и предотвращающий образование здесь завихрений воздуха. Характерная для каждого вида форма крыла определяется длиной скелетных элементов и второстепенных и первостепенных маховых.
Приспособления к полету отчетливо выражены и в поясе передних конечностей. Мощные коракоиды расширенными нижними концами прочно соединяются малоподвижными суставами передним концом грудины. Узкие и длинные лопатки срастаются со свободными концами коракоидов, образуя глубокую суставную впадину для головки плеча. Крепость костей плечевого пояса и их прочное соединение с грудиной обеспечивает крыльям опору в полете. Удлинение коракоидов увеличивает площадь прикрепления мышц крыла и выносит вперед на уровень шейных позвонков, плечевой сустав; это позволяет укладывать крыло сбоку туловища в покое и выгодно аэродинамически, ибо в полете центр тяжести птицы оказывается на линии, соединяющей центры площадей крыльев (обеспечивается устойчивость). Ключицы срастаются в вилочку, расположенную между свободными концами коракоидов и выполняющую роль амортизатора, смягчающего толчки при взмахах крыла.
Задние конечности и тазовой пояс испытывают преобразования, связанные с тем, что при движении по суше на них переносится вся тяжесть тела. Скелет задней конечности образован мощными трубчатыми костями. Общая длина ноги даже у «коротконогих» видов превышает длину туловища. Проксимальный конец бедра заканчивается сочленяющейся с тазом округлой головкой, а на дистальном конце рельефные поверхности образуют с костями голени коленный сустав. Его укрепляет лежащая в мускульном сухожилии коленная чашечка. Основной элемент голени — костный комплекс, который можно назвать голено-предплюсна, или тибиотарзус, так как к хорошо развитой большой берцовой кости прирастает, образуя ее дистальный конец, верхний ряд косточек предплюсны. Малая берцовая кость сильно редуцирована и прирастает к верхней части наружной поверхности большой берцовой кости. Ее редукция связана с тем, что у большинства птиц все элементы конечности двигаются в одной плоскости, вращательные движения в дистальной части конечности ограничены.
Дистальный (нижний) ряд косточек предплюсны и все кости плюсны сливаются в единую кость — цевку, или плюсно-предплюсну; появляется добавочный рычаг, увеличивающий длину шага. Так как подвижный сустав располагается между двумя рядами элементов предплюсны (между костями, слившимися с большой берцовой костью, и элементами, вошедшими в состав цевки), то его, как и у пресмыкающихся, называют интертарзальным. К дистальному концу цевки прикрепляются фаланги пальцев.
Как и у всех наземных позвоночных, тазовый пояс птиц образован срастающимися тремя парами костей. Широкая и длинная подвздошная кость срастается со сложным крестцом. К ее наружному краю прирастает седалищная кость, с которой срастается палочковидная лобковая кость. Все три кости участвуют в образовании вертлужной впадины, в которую входит, образуя тазобедренный сустав, головка бедра. Лобковые и седалищные кости у птиц не срастаются друг с другом по средней линии тела; такой таз называют открытым. Он дает возможность откладывать крупные яйца и, может быть, способствует интенсификации дыхания, не ограничивая подвижности брюшной стенки в тазовой области.
Большая поверхность таза и его прочное соединение с осевым скелетом обеспечивают опору задним конечностям и создают возможности для прикрепления мощных ножных мышц. Длинные крепкие кости конечностей, резкий рельеф их суставных поверхностей наряду с хорошо развитой и дифференцированной ножной мускулатурой обеспечивают интенсивное движение в разнообразных условиях.
Мускулатура птиц дифференцирована сильнее, а ее относительная масса больше, чем у пресмыкающихся. Это связано, с большей подвижностью птиц и разнообразием их движений. Компактность тела, обусловленная требованиями аэродинамики, в значительной степени достигается тем, что наиболее мощные мышцы, осуществляющие движения конечностей, расположены на туловище, а их сухожилия идут к конечностям. Сильное развитие связок укрепляет соединение отдельных элементов скелета. Очень сложна мускулатура шеи, обеспечивающая высокую подвижность головы, что важно и при схватывании добычи, и при ориентировке, и в полете.
Из мышц передней конечности в первую очередь заслуживают упоминания две. Подключичная мышца прикрепляется на коракоиде, теле и гребне грудины, а ее сухожилие оканчивается на головке плеча; сокращение этой мышцы поднимает крыло. Над ней лежит, прикрепляясь к грудине и ее килю, к коракоиду и коракоидно-ключичной связке, большая грудная мышца, опускающая в полете крыло; ее сухожилие прикрепляется тоже к головке плеча. Обе большие грудные мышцы составляют от 10 до 25% от общей массы птицы и превышают массу подключичных мышц в 3-20 раз. Особенно велики эти мышцы у птиц, летающих стремительным, маневренным полетом. Кроме этого, работой крыла в полете управляют еще несколько десятков более мелких мышц, располагающихся на туловище, плече и предплечье.
Движения задних конечностей осуществляют более 30 мышц. Более крупные из них широкими основаниями прикрепляются к костям таза, мелкие мышцы расположены на бедре и голени. Расположенный на задней поверхности голени глубокий сгибатель пальцев образует сухожилие, проходящее по задней стороне интертарзального сустава и цевки, а затем разделяющееся на четыре ветви и заканчивающееся на нижней поверхности концевых фаланг пальцев. Поверхность концевых сухожилий и дно широких соединительнотканных влагалищ, по которым они двигаются, имеют поперечную ребристость. Когда птица садится на ветку и сжимает пальцы, под давлением ее массы сухожилия прижимаются к стенке влагалищ, и их ребрышки сцепляются: пальцы остаются в сжатом состоянии и при расслаблении мускула. Чтобы разошелся этот «Автоматический замок», нужно сокращение мышц — разгибателей пальцев. Это дает возможность птицам спать, сидя на ветви, с расслабленной мускулатурой.
Дыхательные движения грудной клетки осуществляются при помощи межреберных и других мышц стенок тела. Несколькими мышцами осуществляется движение хвоста. По сравнению с пресмыкающимися, у птиц лучше развита подкожная мускулатура, позволяющая менять положение перьев на больших участках тела. Мелкие мускульные пучки кориума меняют положение отдельных перьев.
Для птиц характерно накопление в мышцах миоглобина, позволяющего создавать резервный запас кислорода, утилизируемый в период интенсивной работы. Наивысшая концентрация миоглобина отмечена в большой грудной мышце, мускулатуре мускульного желудка и сердца. Концентрация мышечного гемоглобина выше у птиц, летающих активным полетом, у ныряющих птиц и птиц высокогорий. Во всех случаях, однако, концентрация гемоглобина в крови выше, чем в мышцах.
Лазанье и прыжки в ветвях при помощи задних и отчасти передних конечностей и использование крыльев для планирования — таковы, видимо, основные способы движения примитивных древних птиц. И сейчас большинство видов в той или иной степени связана с кронами деревьев и кустов. В ветвях птицы обычно передвигаются прыжками, иногда помогая одиночными взмахами крыльев. Обычный для них тип строения лапы — три пальца вперед, один назад — позволяет прочно охватывать ветки. Дальнейшая специализация к древесному образу жизни часто сопровождается изменением строения лапы — два пальца направлены вперед, два назад, что, вероятно, еще более облегчает удерживание на ветвях. Прекрасно лазающие в кронах попугаи используют для хватания и мощный клюв. За счет резкого усиления мускулатуры ног, развития сильных пальцев с острыми когтями многие, особенно мелкие, виды птиц освоили лазанье по вертикальным стволам (поползни, пищухи и др.) и по скалам. Сильные лапы с мощными пальцами и острыми когтями и прочный хвост, служащий добавочной опорой, позволяют дятлам не только лазить по вертикальным стволам, но, удерживаясь на одном месте, долбить кору и древесину.
Многие виды древесных птиц собирают корм и на земле. Мелкие виды при этом, как и в ветвях, прыгают (воробьи и др.), а другие ходят и бегают, попеременно переставляя ноги (белая трясогузка, грач, ворона и др.). Приспособление к наземному образу жизни часто сопровождается укорочением пальцев, особенно направленного назад (иногда он редуцируется), удлинением цевки. Лучшие бегуны среди птиц — потерявшие способность к полету страусоподобные птицы. Трехпалый южноамериканский нанду и двухпалый африканский страус могут бежать со скоростью более 50 км/ч. У обитателей болот и берегов водоемов (цапли, пастушки, кулики и др.) удлинение цевки и голени позволяет бродить по мелководью, не смачивая оперение длинные тонкие пальцы не дают проваливаться на вязком грунте. глухаря, рябчика и других тетеревиных во время осенней линьки, по бокам пальцев вырастают роговые шипики, увеличивающие площадь опоры и уменьшающие проваливание в снег, а у белых куропаток на пальцах вырастают длинные перья, превращающие лапу в лыжу — снегоступ. Многие плавающие птицы хорошо «ходят по суше (чайки и др.); у некоторых хороших ныряльщиков ноги далеко отнесены назад и по суше они передвигаться почти не могут (гагары, поганки). Мало и плохо ходят некоторые хорошо летающие виды (ласточки, щурки, стрижи).
Плавание и ныряние
Благодаря малой плотности (оперение, воздушные мешки) могут присаживаться на воду и взлетать с нее: и наземные птицы, например дрозды. Способность к настоящему плаванию, когда птица находится на воде, кормясь и отдыхая, выработалась у представителей разных отрядов. У хороших пловцов тело несколько сплющено в спинно-брюшном направлении, что придает большую устойчивость на воде, сильно развита пневматичность скелета, оперение более плотное, чем у наземных птиц, обильнее пух. При плавании ноги несколько относятся назад, а сила гребка увеличивается тем, что пальцы соединены плавательной перепонкой (все четыре — у пеликанообразных и только три, направленные вперед, — у трубконосых, гагар, чаек, гусеобразных) или каждый палец окаймляет кожистая оторочка (поганки, лысухи, кулики плавунчики).
Ныряние для птиц затруднено из-за их низкой плотности. Несмотря на это некоторые виды успешно добывают пищу в толще воды. Ныряющих птиц можно разделить на две группы.
К первой относятся виды, ныряющие с пикирования, когда за счет инерции птица погружается в воду на небольшую глубину (на 1-2 длины тела), схватывая в этот момент добычу, а затем выбрасывается, как пробка, наружу. Так ныряют как хорошо плавающие птицы (некоторые пеликанообразные, трубконосые, чайки), так и птицы, не умеющие плавать, например скопа и орлан белохвост из соколообразных, зимородки из ракшеобразных и др. Приспособлений к нырянию у них мало: немного плотнее оперение, высокая скорость пикирования и т. п.
Вторая группа — настоящие ныряющие птицы, ныряя, погружаются на большую глубину (некоторые виды — на несколько десятков метров) и активно двигаются в толще воды, отыскивая и преследуя добычу. У таких видов отчетливее выражены приспособления к нырянию. Уменьшается пневматичность костей. Повышается плотность, (до 0,7-0,8); прижимая оперение и произвольно уменьшая объем воздушных мешков в момент ныряния, птицы способны повысить плотность почти до 1,0. Укорочение крыльев улучшает обтекаемость тела в воде, но сопровождается ухудшением полета. Бедренный сустав сдвигается назад. Поэтому на суше тело приобретает более или менее вертикальное положение (бакланы, чистиковые, особенно пингвины), походка делается неуклюжей. Движение в толще воды совершается с помощью гребных движений ног, делающих резкие толчки назад. Чистиковые птицы в толще воды работают полурасправленными крыльями («подводный полет»), а вытянутые назад ноги с расправленными пальцами выполняют лишь роль рулей. У пингвинов при плавании на поверхности воды и при нырянии функцию движителя выполняют преобразованные в ласты крылья, а ноги работают лишь как рули.
Продолжительность ныряния не превышает 2-3 мин, лишь у немногих видов (пингвины) — до 5-7 мин. У ныряющих птиц выше резервы кислорода: больше гемоглобина в крови и в мышцах. В момент погружения уменьшается число сердечных сокращений, а кровью продолжает интенсивно снабжаться лишь головной мозг и некоторые внутренние органы (кишечник, почки).
Работающие мышцы используют кислород, связанный мышечным гемоглобином, и переходят на анаэробные процессы (гликолиз), о чем свидетельствует резкое повышение концентрации молочной кислоты после ныряния в мышцах и в крови. В толще воды обычно двигаются со скоростью 1-2 м/с; пингвины способны развивать скорость до 8-10 м/с (до 36 км/ч). Наиболее специфическая форма движения птиц, определившая основные черты организации этого класса, — полет. Аэродинамика полета птиц сложна и до сих пор известна лишь в общих чертах. Связано это с тем, что в полете меняется положение маховых и площадь крыла, с разной скоростью и под разными углами к горизонту двигаются кистевая и основная части крыла и т. п.
Грубо схематично физическую основу полета можно характеризовать так. Крыло всегда сверху более или менее выпукло, а снизу вогнуто, передний край — крыла более толстый (здесь лежит скелет, мышцы и несколько слоев перьев), задний — тонкий и эластичный (образован лишь вершинами маховых). Обтекая верхнюю, выпуклую поверхность крыла, встречный поток воздуха ускоряет движение, и над крылом образуется область пониженного давления. Создается подъемная сила, подсасывающая (поднимающая) крыло вверх. Когда в полете птица опускает крыло, одновременно протекают два процесса. Отгибающиеся под давлением воздуха первостепенные маховые концевой части крыла создают пропеллирующий эффект, в результате чего возникает тяговая сила, толкающая крыло (и птицу) вперед. Одновременно воздух обтекает основную часть крыла (область второстепенных маховых) и здесь создается, за счет разности давлений над крылом и под крылом, подъемная сила, преодолевающая силу тяжести птицы. При подъеме крыла вверх маховые несколько поворачиваются, пропуская воздух, и поэтому подъем совершается с меньшим усилием. Двигающаяся вверх и назад вершина крыла создает некоторую дополнительную силу тяги, а основная часть крыла по-прежнему создает подъемную силу. Подъемная сила возникает и при обтекании воздухом тела и хвоста летящей птицы.
Такой полет, когда птица ритмично поднимает и опускает крылья, называется машущим. Изменяя площадь крыла и его наклон («угол атаки»), варьируя частоту взмахов, птица изменяет величину тяги и подъемной силы, меняя тем самым скорость и высоту полета. Различия в размерах и форме тела, размерах и форме крыльевой хвоста, в интенсивности и амплитуде взмахов крыла определяют свойственный каждому виду характер полета. Медленный, со спокойными и редкими взмахами крыльев полет цапли отличен от стремительного и маневренного полета ласточек и стрижей и от быстрого, но прямолинейного полета уток. Но все эти птицы летают машущим полетом. Одна из форм машущего полета — трепещущий полет, когда птица, усиленно работая крыльями, на короткое время «повисает» в воздухе на одном месте. Так делают чайки, крачки, мелкие хищники, высматривающие добычу. Сходным образом «повисают» в воздухе около цветка высасывающие нектар колибри; при этом крыло совершает 50-80 взмахов в секунду.

Второй тип полета — парящий;птица с распростертыми практически неподвижными крыльями перемещается, используя энергию потоков воздуха. Различают статическое и динамическое парение. Статическое парение возможно над материками, где устойчивые восходящие токи воздуха возникают на стыках ландшафтов (лес и поле и т. п.) или при обтекании воздухом препятствий — обрывов, горных вершин. Для птиц, использующих устойчивые токи воздуха, характерны большие, широкие, закругленные крылья с расходящимися на концах вершинами первостепенных маховых. Такой полет используют хищные птицы, аисты, пеликаны. Широкими кругами птицы постепенно набирают высоту и затем кружат, высматривая добычу, или, планируя с потерей высоты, перемещаются в нужном направлении. Динамическое парение свойственно морским птицам (альбатросы, буревестники, чайки), имеющим длинные, но узкие, с заостренной вершиной крылья. Используя завихрения воздуха над волнами или разную скорость воздушных, потоков, птица по ветру планирует вниз, набирая скорость, и у самой воды, где скорость ветра замедлена трением о воду, поворачивается против ветра и взмывает вверх, где воздух движется быстрее. Так птица может парить часами, высматривая добычу и хватая ее с пикирования. При отсутствии ветра эти птицы парить не могут и, плавая, пережидают безветрие.
Летающие планирующим полетом птицы способны и к машущему полету. К нему они прибегают, чтобы найти восходящий термический поток, подлететь к гнезду, увернуться от опасности и т. п. Однако долго летать машущим полетом они не могут. С другой стороны, птицы, летающие машущим полетом, временами переходят на скольжение или планирование. В общем, каждый вид пользуется характерным для него полетом, но при необходимости способен менять как характер полета, так и его скорость. Лесные мелкие воробьиные птицы летают со скоростью 25-40 км/ч, голуби — 30-60, утки и многие кулики — 65-80 км/ч; ловящие насекомых в воздухе ласточки обычно летают со скоростью 40-60 км/ч, а стрижи — даже до 100- 120 км/ч. Самый быстрый летун нашей фауны — живущий в Приморье колючехвостый стриж развивает скорость до 170 км в час. Таковы скорости, с которыми птицы могут летать длительное время. Например, почтовый голубь, летя со скоростью 50- 60 км/ч, способен пролететь без остановок около 600 км. На короткое время птицы способны развивать и большие скорости. Крупные соколы летают со скоростью 60-70 км/ч, но при броске за добычей развивают скорость более 100 км/ч, а бросаясь на нее с высоты, за счет инерции пикирования — и до 300-350 км/ч. Такую же скорость, но на очень короткое время — на десятки секунд — способны развивать увертывающиеся от хищника стрижи, голуби, утки.
Лётные возможности и характер полета каждого вида тесно связаны со всеми особенностями его жизни: предпочитаемыми местообитаниями, набором кормов и способами его добывания, приемами избегания опасности, дальностью сезонных перемещений и т. п. Виды, кормящиеся в воде и при опасности прячущиеся в зарослях или ныряющие, обладают быстрым, но не маневренным полетом (утки, гагары и др.). Кормящиеся на земле и при опасности старающиеся сначала затаиться куриные птицы имеют короткие и широкие крылья, позволяющие им с шумом стремительно взлетать под носом ошеломленного хищника и, пролетев небольшое расстояние, скрыться в зарослях; громкое хлопанье крыльев при взлете предупреждает об опасности соседей. Быстрый полет рябков позволяет им гнездиться вдали от водоемов и ежедневно летать на водопой за десятки километров. Разыскивая добычу, орлы, канюки, коршуны часами парят в воздухе над открытыми пространствами. Подстерегающие добычу и преследующие ее в лесу ястребы обладают маневренным полетом, позволяющим преследовать добычу в кронах деревьев. Разыскивающие добычу на земле и в поверхностных слоях воды чайки и крачки чередуют маневренный машущий полет с динамическим парением и кратковременным «зависанием». Часами носятся в воздухе, ловя мелких насекомых, стрижи; они чередуют быстрый машущий полет со скольжением. Эти быстрые и неутомимые летуны пьют на лету, спариваются в воздухе и даже способны ночью спать, медленно, со скоростью 30-40 км/ч, летая кругами на большой высоте.
Литература: Зоология позвоночных. Ч. 2. Пресмыкающиеся, птицы, млекопитающие. Наумов Н. П., Карташев Н. Н.
Источник



