
- Пили ворсинки фимбрии локализация типы биологическая роль способ обнаружения
- Значение пилей в жизни бактерий
- Внешний вид фимбрий
- Для чего прокариотам пили
- Разные типы ворсинок
- Пили 1 типа
- Ворсинки 2 типа
- Половые фимбрии (3 тип)
- Участие F-пили в процессе передачи информации
- Специфическая направленность фимбрий 4 типа
Пили ворсинки фимбрии локализация типы биологическая роль способ обнаружения
• Пили представляют собой внеклеточные белковые структуры, которые осуществляют самые разнообразные функции, включая обмен ДНК адгезию и образование биофильма клетками прокариот
• Многие адгезивные пили собираются с участием системы шаперон-Usher-белок. Сборка происходит на наружной мембране с участием Usher-белка, образующего пору, сквозь которую проходят субъединицы пили, и шаперона периплазмы, способствующего их скручиванию и прохождению через пору
• Жгутики представляют собой внешние структуры клетки, которые служат пропеллерами, обеспечивающими ее движение
• У прокариот жгутики состоят из множественных сегментов, каждый из которых образуется при сборке белковых субъединиц
От поверхности прокариотической клетки отходят два типа придаточных структур, пили и жгутики. Пили представляют собой нитевидные олигомеры белков, присутствующие на клеточной поверхности. Существуют различные типы пилей. Например, F-пили участвуют в клеточной конъюгации и в переносе ДНК. Когда эти придаточные структуры были впервые обнаружены, их назвали «фимбрии» (лат. fimbria — нить, волокно). Их присутствие коррелировало со способностью Е. coli агглютинировать красные кровяные клетки.
Позже для обозначения фибриллярных структур (F-пили), связанных с процессом переноса генетического материала между организмами при конъюгации, был предложен термин пили (или пилюс) (лат. pilus — волос). С тех пор этот термин стал общим для описания всех типов ворсинчатых придаточных структур, и используется наряду с термином фимбрия.
Взаимодействие клеток бактерий с другими прокариотическими и эукариотическими клетками с участием ворсинок часто служит важным этапом заселения эпителия, проникновения микробов в клетки хозяина, обмена ДНК и формирования биопленок. Пили могут служить рецепторами бактериофагов. Основная функция большинства пилей состоит в структурном обеспечении позиционирования специфических молекул, участвующих в клеточной адгезии. Адгезивные субъединицы ворсинок (адгезины) представляют собой минорные компоненты их кончиков, однако основные структурные субъединицы также могут функционировать в качестве адгезинов.
Часто адгезивные пили представляют собой важные факторы заселения микробами организма хозяина. Например, при инфекциях мочевых путей патогенными бактериями Е. coli, клетки прикрепляются к эпителию мочевого пузыря с помощью пилей типа I. Пили этого типа присутствуют у многих грамотрицательных микроорганизмов. Они представляют собой сложные структуры, состоящие из толстого тела, соединенного с тонким фибриллярным концом. На конце расположены молекулы адгезина FimH, которые связываются с остатками маннозы на поверхности клеток хозяина.
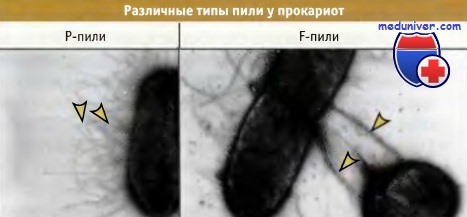 Два типа пили у клеток прокариот.
Два типа пили у клеток прокариот.
Р-пили короче, чем F-пили, и принимают участие в адгезии клеток.
F-пили участвуют в конъюгации и в переносе ДНК между клетками.
Фотографии любезно предоставлены Мэтт Чэпмен (слева) и Роном Скарри (справа), биологический факультет Сиднейский университет.
Сборка пилей представляет собой сложный процесс, в котором участвуют структурные белки, составляющие тело пили, и дополнительные белки, способствующие сборке субъединиц на поверхности клетки. Все структурные компоненты, необходимые для процесса сборки пилей на поверхности грамотрицательных микроорганизмов, должны транслоцироваться через цитоплазматическую мембрану в периплазму и далее, через внешнюю мембрану. В завершении процесса сборки участвуют два специфических белка: шаперон, присутствующий в периплазме, и транспортный белок внешней мембраны, который называется Usher-белок.
Процессы, в которых функционируют эти белки, обеспечивают биогенез более 30 различных типов ворсинчатых структур. Как показано на рисунке ниже, комплексы шаперонов с субъединицами образуются в периплазме и на наружной мембране взаимодействуют с Usher-белком, где высвобождается шаперон. При этом на субъединицах открываются интерактивные поверхности, что обеспечивает их дальнейшую сборку в пили. Исследования пилей типа I и Р показали, что адгезин-шапероновые комплексы (PapDG или FimCH) обладают большим сродством к Usher-белку, и адгезины представляют собой начальные субъединицы, которые собираются в пили.
Включение остальных субъединиц отчасти определяется кинетикой образования на Usher-белке комплекса с шапероном. Наряду с функционированием в качестве сборочной платформы, Usher-белок, вероятно, играет также и другие роли в сборке ворсинок. По данным электронной микросокопии высокого разрешения, PapС Usher имеет вид кольцевых комплексов диаметром 15 нм, которые в середине имеют пору размером 2 нм. После отщепления от шаперона, которое происходит на Usher-белке, субъединицы включаются в растущую структуру пили, которая, как считают, должна выталкиваться через центральную пору комплекса в виде толстой линейной фибриллы, состоящей из одной субъединицы.
Большинство микроорганизмов обладает подвижностью, и часто она обеспечивается длинными структурными придатками, которые называются жгутиками. У грамположительных и грамотрицательных бактерий жгутики собираются на поверхности клеток. Когда на полюсе клетки находится один жгутик, такое расположение называется монотрихиальным (или полярным). Если жгутики расположены вокруг клетки, то такое расположение называется перитрихиальным.
Если на одном полюсе клетки находится группа жгутиков, то говорят об их лофотрихиальном расположении (от латинского «хохолок»). Жгутики бактерий отличаются от этих структур эукариотических клеток, которые состоят из микротрубочек и связанных с ними белков и окружены плазматической мембраной.
Жгутики могут быть различной длины, но их диаметр обычно составляет 20 нм. Они не видны в световом микроскопе, если препараты вначале не обрабатывались реагентами, которые увеличивали диаметр жгутиков. На рисунке ниже видно, что жгутики состоят из трех отдельных доменов: филамента, крючка и базального тела. Филамент жгутика состоит из повторяющихся структур флагеллиновых белков. Флагеллины представляют собой высококонсервативные белки бактерий, что позволяет предполагать, что движение клеток с участием жгутиков характерно для примитивных форм живых организмов. В месте присоединения жгутика к клетке находится базальное тело, представляющее собой сложную структуру, состоящую из множества белков.
Филамент жгутика соединяется с базальным телом посредством крючка. У грамотрицательных бактерий базальное тело проходит через наружную мембрану, протеогликан клеточной стенки и цитоплазматическую мембрану. С наружной мембраной жгутик связан посредством L-кольца. Две пары колец, S-М и Р, способствуют прикреплению жгутика к цитоплазматической мембране и к клеточной стенке соответственно. Каждое кольцо состоит из множества мембранных белков. На цитоплазматической мембране находятся два белка Mot, которые выполняют роль моторов, приводящих жгутики в движение. Еще один набор белков встроен в цитоплазматическую мембрану и выполняет реверсную функцию по отношению к моторам жгутика. Поскольку у грамположительных организмов наружная мембрана отсутствует, у них есть только S-М кольца.
В образовании и сборке филаментов жгутиков участвует несколько десятков различных генов. Их активность строго регулируется в соответствии с порядком процесса сборки. Так, первыми экспрессируются гены, участвующие в сборке базального тела и крючка, а затем наступает очередь генов, ответственных за образование субъединиц жгутика. Экспрессии флагеллиновых субъединиц не происходит до тех пор, пока не завершилась сборка крючка. В этот момент через канал крючка выходит супрессор транскрипции, и, таким образом, снимается подавление экспрессии флагеллина. Субъединицы флагеллина экспортируются через жгутик и добавляются к его растущему концу.
Такой механизм обеспечивает сборку филамента только после образования структуры крючка. Эта структура также имеет отношение к другим секреторным системам белков.
Система хемотаксиса определяет наличие питательных компонентов и затем определяет направление вращения жгутика. В отсутствие питательных компонентов, жгутики вращаются по часовой стрелке, что вызывает поворот клетки. Движение клетки по направлению к молекулам химического соединения или от них называется хемотаксис. В данном разделе мы рассмотрим движение прокариотической клетки в присутствии аттрактанта, являющегося питательным продуктом.
Для того чтобы обеспечить клетке такое движение, жесткий жгутик должен вращаться подобно пропеллеру, за счет энергии, доставляемой протонной движущей силой. Движение клетки состоит из серии прямых пробегов, за которыми следуют ее быстрые беспорядочные повороты. Когда жгутики вращаются против часовой стрелки, клетка перемещается по прямой линии, а при вращении по часовой стрелке клетка совершает повороты. Поскольку в результате поворотов клетка занимает случайные позиции, можно было бы думать, что общий итог движения окажется нулевым. Однако периодичность пробегов регулируется в соответствии с доступностью питательного компонента: более длинные пробеги характерны для движения клетки по направлению к источнику питания, и количество поворотов возрастает, когда клетка направляется от него.
Хотя направление отдельных пробегов все еще случайно, общий результат проявляется в движении клетки в сторону аттрактанта.
Пути передачи сигнала хемотаксиса у прокариот характеризуются чрезвычайно консервативной природой. Единственным из известных организмов, в геноме которого отсутствуют гены хемотаксиса, является Mycoplasma. Практически у всех прокариот обнаружены следующие консервативные белки хемотаксиса: CheR, CheA, CheY, CheW, и CheB. При протекании сложного каскада событий, включающих фосфорилирование и метилирование, эти белки обеспечивают сложный, скоординированный и высокогибкий ответ клетки на присутствие аттрактантов и репеллентов в окружающей среде. Мы опишем, как происходят эти события в клетках Е. coli.
Присутствующие в окружающей среде аттрактанты или репелленты связываются с рецепторами, расположенными на цитоплазматической мембране. С этими рецепторами взаимодействует киназа CheA, также расположенная в цитоплазматической мембране. Эта киназа фосфорилирует CheY, который затем связывается с мотором жгутика, что приводит к переключению направления его вращения и к повороту клетки. Под действием фосфатазы CheZ из CheY удаляется фосфатная группа. При низкой концентрации аттрактанта происходит аутофосфорилирование CheA, фосфатная группа переносится на CheY, и последний мигрирует к мотору жгутика, изменяя характер движения клетки на поворот.
Система хемотаксиса характеризуется еще одним уровнем сложности, который позволяет клетке постоянно адаптироваться к условиям, существующим в окружающей среде. По мере своего продвижения по градиенту концентрации химических соединений, клетка может реагировать на возникающие небольшие флук-туации. Такая кратковременная память обеспечивается за счет метилирования мембранных рецепторов. CheR метилирует мембранные рецепторы, a CheB удаляет метальные группы.
Метилирование рецепторов увеличивает активность киназы CheA, что приводит к десенсибилизации системы. В свою очередь, CheB также фосфорилируется CheA; это вызывает увеличение метилэстеразной активности CheB и замыкает цикл обратной связи для сигнального каскада.
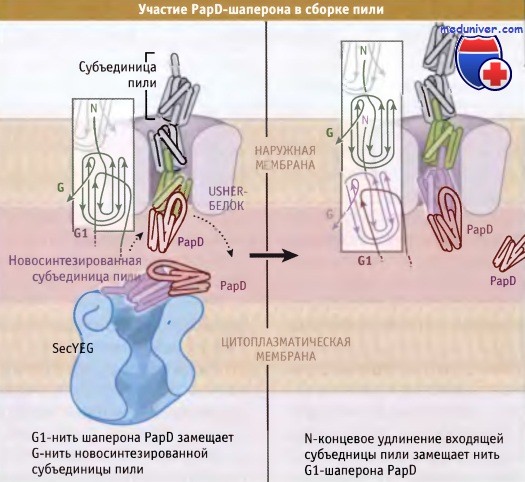 Для сборки пили необходим шаперон PapD и мембранный Usher-белок.
Для сборки пили необходим шаперон PapD и мембранный Usher-белок.
Слева: нить G1 шаперона PapD завершает формирование нативной структуры новосинтезированной субъединицы пили,
располагаясь между двумя нитями этой субъединицы.
Справа: в процессе сборки пили N-концевая нить новосинтезированной субъединицы замещает G1-цепь шаперона PapD,
связанную с предыдущей субъединицей пили. 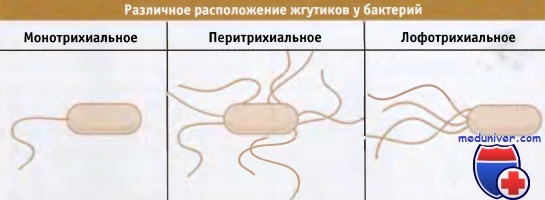 У различных типов бактерий жгутики расположены по-разному.
У различных типов бактерий жгутики расположены по-разному. 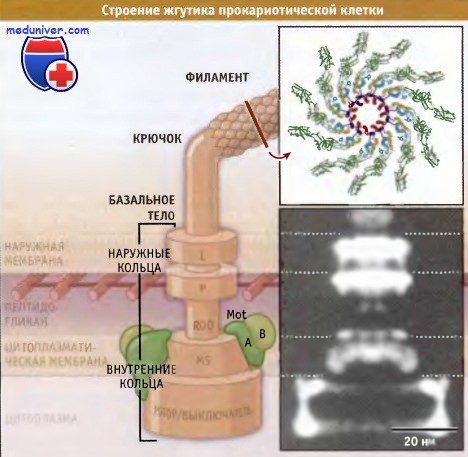 Жгутики прокариот состоят из трех сегментов, каждый из которых содержит многочисленные белки.
Жгутики прокариот состоят из трех сегментов, каждый из которых содержит многочисленные белки.
На врезке вверху представлено изображение структуры жгутика, полученное с помощью электронного микроскопа,
а внизу — электронная микрофотография области базального тела и крючка.
Филамент жгутика состоит из флагеллиновых субъединиц, которые собраны в спиральную структуру, содержащую 11 субъединиц на виток. 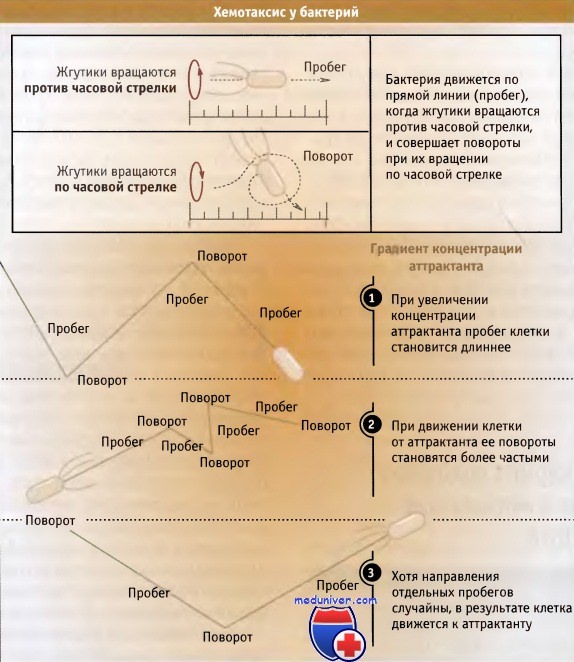 Движение бактерии по направлению к молекулам питательных веществ
Движение бактерии по направлению к молекулам питательных веществ
и от них можно представить в виде длительных прямолинейных треков и, соответственно, большим числом поворотов.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
Значение пилей в жизни бактерий
Наряду со жгутиками прокариоты могут обладать и другими внеклеточными образованиями. В середине ХХ века было установлено, что бактерии способны формировать специфическую группу поверхностных образований. Их называли ворсинками, ресничками, фимбриями. Сегодня их называют пилями бактерий.

Внешний вид фимбрий
Внешне пили, или фимбрии, как их называли до 1956 г., выглядят как микроскопические волоски, покрывающие клетку бактерии. На 1 клетку прокариота может приходиться от нескольких единиц до тысяч ворсинок.
Хотя они, как и жгутики, являются поверхностными образованиями, однако между собой имеют больше различий, чем сходств.
По размеру пили намного меньше жгутиков, в среднем в 3 раза тоньше (не более 10 нм), и длиной не превышают 1,5 мкм.
По строению, несмотря на то, что как пили, так и жгутики состоят из белковых клеток, они также различаются:
- пили, или фимбрии, представляют собой легкую цепочку проводящих белков цилиндрической формы, отходящую от поверхностного слоя клетки;
- жгутики являются более громоздкими по строению, с наличием сложных структур (стержень, базальное тело, кольца и другое).
Столь явное различие в строении поверхностных образований прокариотов связано с совершенно разными задачами, которые они решают в процессе жизнедеятельности бактериальной клетки.
Для чего прокариотам пили
К примеру, если жгутики бактерий обеспечивают возможность передвигаться, то фимбрии не имеют никакого отношения к перемещению в пространстве и присутствуют как у движущихся, так и у неподвижных бактерий.
Неподтвержденным исключением можно считать участие пилей 4 типа в скользящем движении прокариотов всей колонией, характерном для многих протеобактерий (и миксобактерий), скользящих бактерий, циано-, несерных бактерий и других.
В отличие от жгутиков функции пилей бактерий изучены достаточно слабо, но совершенно очевидно, что одной из них является способность обеспечить прикрепление клетки бактерии к питательному субстрату.
Разные типы ворсинок
Пили не являются однородными образованиями, их различают как минимум 4 типа, каждый из которых выполняет свои функции, причем одна клетка может являться носителем несколько разных типов фимбрий.
Пили 1 типа
Фимбрии бактерий 1 типа образуются из пилина (белок) и отличаются крайне прочной связью с прокариотом. Чтобы отделить такую фимбрию от бактериальной клетки, требуются усилия, многократно превышающие необходимое воздействие для отделения половых пилей или жгутиков.
Для пилей 1 типа характерно расположение перитрихиально – по всей поверхности бактерии.
Исследования методами выявления свойств показали, что пили 1 типа являются химически устойчивыми образованиями – они инертны к растворам щелочей, мочевине и трипсину (фермент, расщепляющий белки).
Разрушаются пили 1 типа при кипячении в растворах с высокой кислотностью, при этом методе воздействия происходит необратимое разрушение (денатурация) белка, образующего фимбрию.
Характерной особенностью пилей 1 типа является:
- способность образовывать пленки и придавать бактериям гидрофобные свойства;
- способность вызывать агглютинацию эритроцитов (выпадение в осадок в результате склеивания) под действием агглютининов.
Основными функциями являются:
- адгезивная – прикрепление бактерий к субстратам;
- защитная – объединение клеток прокариотов, получивших гидрофобные свойства, в группы;
- участие в процессах метаболизма клетки – увеличение всасывающей поверхности.
Ворсинки 2 типа
Эта группа имеет очень много общего с предыдущей, однако не обладает характерными особенностями 1 типа – пили не участвуют в формировании пленок и не приклеиваются к эритроцитам (агглютинация), провоцируя выпадение их в осадок.
Столь близкое сходство позволяет предположить, что пили 2 типа являются мутантной формой 1 типа.
Половые фимбрии (3 тип)
Современные методы выявления позволили определить, что при горизонтальном переносе генетического материала (конъюгации) ключевую роль играют половые пили.
Возможность непосредственного контакта двух бактериальных клеток с последующей конъюгацией была выявлена в 50-х годах прошлого века в результате исследований двух американских биохимиков – Д.Ледербейга и Э.Тейтема. Данный процесс имеет большое практическое значение, так как позволяет производить обмен наследственными признаками организмам, размножающимся только прямым делением.
Половые фимбрии, их называют F-пили, присутствуют только у бактериальных штаммов, обладающих фактором трансмиссивности – это может быть автономный репликон или его часть.
F-пили представляют собой цилиндрические белковые образования большего диаметра, чем пили 1 или 2 типа, расположенные перпендикулярно к поверхности.
Формирование пиля осуществляется на поверхности цитоплазматической мембраны в точках контакта ее с вешней оболочкой. Сформированная трубочка проходит сквозь слои муреина и внешнюю мембрану.
В случае потери F-пили восстанавливаются – в течение 30 секунд пиль достигает половины своей величины. Для формирования полноценной трубочки необходимо от 4 до 5 минут.
Современные методы выявления позволили определить, что на поверхности бактерии F-пили сохраняются в течение 5 минут, после чего сбрасываются, и процесс повторяется.
F-пили значительно отличаются от ворсинок 1 и 2 типа как по строению, так и по свойствам.
В отличие от последних F-пили легко отделяются от бактериальной клетки даже при незначительном встряхивании.
Методами физико-химического анализа было определено, что в составе F-пили отсутствует целый ряд α-аминокислот, характерных для белка пилей 1 типа, но через ковалентную связь присоединены остаток D-глюкозы и две фосфатные группы.
В связи с иной химической структурой на F-пили не адсорбируются обычные фаги, а только специфичные для них, называемые мужскими фагами.
Участие F-пили в процессе передачи информации
Процесс передачи части генетической информации подразумевает наличие пары клетка-донор и клетка-реципиент.
- Первоначально клетка-донор формирует F-пиль.
- F-пиль донора фиксируется на клетке-реципиенте.
- В F-плазмиде донорной клетки осуществляется разрыв одной нити ДНК, которая передается реципиенту.
- Обе бактерии достраивают вторую цепочку ДНК и восстанавливают F- плазмиду. Клетка-реципиент превращается в донора.
Микроскопические методы исследований позволили определить, что образование F-пилей характерно только для растущих и активных клеток, при переходе в стационарную фазу роста бактерии теряют свою способность образовывать половые пили и становятся плохими донорами.
Специфическая направленность фимбрий 4 типа
Пили 4 типа принимают участие в обеспечении разновидности скользящего движения бактерий всей колонией.
Сам процесс скольжения с участием пилей 4 группы предполагает наличие 2 систем движения:
- А-система — секретирует слизь на полюсе направления движения микроорганизма.
- S-система – роение; обеспечивается последовательным сокращением и удлинением пилей 4 типа, подобное подтягиванию.
Механизм данного вида бактериального движения на сегодняшний день находится в процессе изучения, и большинство выводов носят предположительный характер.
Образование высшее филологическое. В копирайтинге с 2012 г., также занимаюсь редактированием/размещением статей. Увлечения — психология и кулинария.
Источник



