
- Гаструляция
- Типы гаструляции
- Процесс и способы гаструляции
- Перечислите особенности гаструляции у плацентарных млекопитающих и человека.
- Первая фаза гаструляции у млекопитающих осуществляется способом
- Гаструляция
- Гаструляция млекопитающих
- Развитие млекопитающих
- Гаструляция у млекопитающих
- Гаструляция у млекопитающих и человека
- Внезародышевые органы млекопитающих и типы плацент
- Типы плацент.
Гаструляция

Типы гаструляции
При гаструляции продолжаются изменения, начавшиеся на стадии бластулы, и поэтому разным типам бластул соответствуют и различные типы гаструляции. Переход из бластулы в гаструлу может осуществляться 4-мя основными способами: инвагинацией, иммиграцией, деляминацией и эпиболией.
Инвагинация или впячивание наблюдается в случае целобластулы. Это наиболее простой способ гаструляции, при котором вегетативная часть впячивается в бластоцель. Вначале появляется небольшое углубление в вегетативном полюсе бластулы. Затем клетки вегетативного полюса все больше и больше впячиваются в полость бластоцеля. В последующем эти клетки доходят до внутренней стороны анимального полюса. Первичная полость, бластоцель, при этом вытесняется и видна только с двух сторон гаструлы в местах изгиба клеток. Зародыш принимает куполообразную форму и становится двухслойным. Его стенка состоит из наружного листка – эктодермы и внутреннего – энтодермы. В результате гаструляции образуется новая полость – гастроцель или полость первичной кишки. Она сообщается с внешней средой с помощью кольцеобразного отверстия – бластопора или первичного рта. Края бластопора называются губами. Различают спинную, брюшную и две боковых губы бластопора.
По последующей судьбе бластопора всех животных разделяют на две большие группы: первично- и вторичноротых. К первичноротым относят животных, у которых бластопор остается постоянным или дефинитивным ртом у взрослой особи (черви, моллюски, членистоногие). У других животных (иглокожие, хордовые) бластопор или превращается в заднепроходное отверстие, или зарастает, а ротовое отверстие возникает заново на переднем конце тела зародыша. Таких животных называют вторичноротыми.
Иммиграция или вселение является наиболее примитивной формой гаструляции. При этом способе происходит перемещение отдельных клеток или группы клеток из бластодермы в бластоцель с образованием энтодермы. Если вселение клеток в бластоцель происходит лишь со стороны одного полюса бластулы, то такая иммиграция называется униполярной, а с различных участков бластулы – мультиполярной. Униполярная иммиграция свойственна некоторым гидроидным полипам, медузам и гидромедузам. В то время, как мультиполярная иммиграция является более редким явлением и наблюдается у некоторых гидромедуз. При иммиграции внутренний зародышевый листок – энтодерма может образовываться сразу в процессе проникновения клеток в полость бластоцеля. В других случаях клетки могут заполнять полость сплошной массой, а затем выстраиваться упорядоченно возле эктодермы и образовывать энтодерму. В последнем случае гастроцель появляется позднее.
Деляминация или расслаивание сводится к расщеплению стенки бластулы. Клетки, которые отделяются внутрь, образуют энтодерму, а наружные – эктодерму. Такой способ гаструляции наблюдается у многих беспозвоночных и высших позвоночных животных.
У некоторых животных в связи с увеличением количества желтка в яйцеклетке и уменьшением полости бластоцеля гаструляция только путем инвагинации становится невозможной. Тогда гаструляция происходит способом эпиболии или обрастания. Этот способ состоит в том, что мелкие анимальные клетки усиленно делятся и обрастают вокруг более крупных вегетативных. Мелкие клетки образуют эктодерму, а клетки вегетативного полюса формируют энтодерму. Такой способ гаструляции наблюдается у круглоротых и земноводных.
Процесс и способы гаструляции
Однако, все описанные способы гаструляции редко встречаются отдельно, обычно они комбинируются. Например, вместе с обрастанием может происходить впячивание (земноводные). Расслаивание может наблюдаться вместе с инвагинацией и иммиграцией (рептилии, птицы и др.).
Следовательно, в процессе гаструляции часть клеток из наружного слоя бластулы перемещается внутрь. Это вызвано тем, что в процессе исторического развития одни клетки приспособились к развитию в непосредственной связи с внешней средой, а другие – внутри организма.
На причины гаструляции единого взгляда не существует. Согласно одним взглядам, гаструляция происходит благодаря неравномерному росту клеток в разных частях зародыша. Румблер (1902) объяснял процесс гаструляции изменением формы клеток внутри и снаружи бластулы. Он считал, что клетки имеют клиновидную форму, внутри бластулы шире, а снаружи уже. Существуют взгляды, что гаструляцию может вызывать резкая интенсивность поглощения воды отдельными клетками. Но наблюдения показывают, что эти различия очень невелики.
Гольтфретер (1943) считал, что анимальный полюс бластулы прикрыт тончайшей пленкой (coat) и поэтому клетки связаны в единую массу. Клетки вегетативного полюса не связаны между собой, имеют бутылковидную форму, удлиняются и втягиваются внутрь. В передвижении клеток может играть роль степень слипания и характер межклеточных пространств. Существует также мнение, что клетки могут передвигаться, благодаря их способности к амебоидному движению и фагоцитозу. Образование третьего зародышевого листка в процессе эмбрионального развития животных осуществляется четырьмя способами: телобластическим, энтероцельным, эктодермальным и смешанным.
У многих беспозвоночных животных (первичноротые) мезодерма образуется из двух клеток – телобластов. Эти клетки обособляются рано, еще на стадии дробления. В процессе гаструляции телобласты располагаются на границе между экто- и энтодермой, начинают активно делиться и образующиеся при этом клетки врастают тяжами между наружным и внутренним листками, образуя мезодерму. Такой способ образования мезодермы называется телобластическим.
При энтероцельном способе мезодерма образуется в виде карманообразных выростов по бокам энтодермы после гаструляции. Эти выпячивания располагаются между экто- и энтодермой, образуя третий зародышевый листок. Такой способ образования мезодермы характерен для иглокожих, ланцетника.
Источник
Перечислите особенности гаструляции у плацентарных млекопитающих и человека.
Несмотря на то, что у млекопитающих отмечаются вторично алецитальные яйцеклетки, а дробление полное асинхронное, в результате чего формируется совершенно отличная от других бластул бластоциста, в фазе гаструляции отмечаются те же процессы, что и у пресмыкающихся и птиц, имеющих полилецитальные яйцеклетки и образующих дискобластулу. Это подтверждает представление о происхождении млекопитающих от предковой группы, у которой яйца были богаты желтком.
Гаструляция у человека протекает в две стадии. Первая стадия (деламинация) приходится на 7-е сут, и происходит в момент имплантации, параллельно с развитием трофобласта, а вторая стадия(иммиграция) — на 14-15-е сут внутриутробного развития.
При деламинации эмбриобласт, образовавший эбриональный диск, расслаивается на два листка: наружный листок — эпибласт и внутренний — гипобласт, обращенный в полость бластоцисты.
Часть клеток эпибласта в дальнейшем образуют стенку амниотического пузырька, который начинает формироваться на 8-е сут. В области дна амниотического пузырька остается небольшая группа клеток эпибласта — материал, который пойдет на развитие тела зародыша и внезародышевых органов.
Второй этап гаструляции млекопитающих и человека – иммиграция.
Перемещение клеток происходит в области дна амниотического пузырька. Асинхронно делящиеся клетки эпибласта образуют скопления клеток – первичную полоску, в головном конце которой возникает гензеновский узелок. В первичной полоске формируется первичная бороздка, через которую начинается выселение клеток в пространство между эпи- и гипобластом. Выселяющиеся клетки формируют листки мезодермы и энтодермы. Часть выселившихся клеток, образовавших энтодерму, внедряется в гипобласт, участвуя в образовании кишечной трубки. В результате зародыш приобретает трехслойное строение в виде плоского диска, состоящего из трех зародышевых листков: эктодермы, мезодермы и энтодермы.
Клетки, выселяющиеся через первичную ямку гензеновского узелка, образуют хорду, которая определяет ось эмбриона.
Дифференцировка зародышевых листков начинается в конце 2-й — начале 3-й нед. Одна часть клеток преобразуется в зачатки тканей и органов зародыша, другая — во внезародышевые органы.
Какие нарушения развития зародыша могут происходить на стадии гаструляции? Каковы их последствия?
ЭКЗОГАСТРУЛЯЦИЯ-нарушение процесса гаструляции у зародышей многоклеточных животных, заключающееся в изменении обычного направления морфогенетич. движений. Происходит при неблагоприятных условиях развития;в эктодерме не дифференцируется нервная ткань, аформируется лишь атипичный эпидермис, тогда как материал хордомезодермы и энтодермы, детерминированный на предыдущих стадиях развития, претерпевает дифференцировку. Явление чётко демонстрирует необходимость контакта между хордомезодермой и эктодермой в период гаструляции у хордовых для детерминации зачатка ЦНС.
Что такое органогенез? Охарактеризовать начальный этап органогенеза- нейруляцию.
Органогенез – процесс образования органов зародыша. Органогенез составляет основное содержимое эмбрионального периода.
Начальной стадией органогенеза является нейруляция.
Нейруляция – процесс образования комплекса осевых органов (нервной трубки, хорды, вторичной кишки).
Нейруляция у всех хордовых животных идет сходным образом. В ответ на индуцирующее действие зачатка хорды (первичного индуктора) недифференцированная спинная эктодерма, расположенная над хордой, преобразуется в нервную пластинку, образованную нейроэпителиальными клетками цилиндрической формы.
Затем края нервной пластинки приподнимаются, формируя боковые валики, ограничивающие нервный желобок. Далее края нервные валики смыкаются друг с другом, образуя нервную трубку с полостью внутри – невроцелем.
Часть нервных клеток, которые не вошли в состав нервной трубки при ее смыкании, образуют нервный гребень. Клетки нервного гребня отличаются способностью к обширным, но строго регулируемым миграциям по всему телу и образуют два главных потока.
Клетки одного из них — поверхностного — включаются в эпидермис или дерму кожи, где дифференцируются в пигментные клетки (меланоциты).
Другая часть клеток мигрирует в брюшном направлении и образует чувствительные спинномозговые ганглии, симпатические нервные узлы, мозговое вещество надпочечников, парасимпатические ганглии. Клетки из черепного отдела нервного гребня дают начало как нервным клеткам, так и ряду других структур, таких, как жаберные хрящи, некоторые покровные кости черепа.
Параллельно с образованием нервной трубки идет завершение формирования хорды, а также вторичной кишки, которая располагается под хордой.
Таким образом, в процессе нейруляции возникает комплекс осевых органов: нервная трубка, хорда, вторичная кишка, представляющих собой характернейшую черту организации тела всех хордовых. Одинаковое происхождение, развитие и взаимное расположение осевых органов выявляют их полную гомологию и эволюционную преемственность.
Дата добавления: 2019-01-14 ; просмотров: 1166 ; Мы поможем в написании вашей работы!
Источник
Первая фаза гаструляции у млекопитающих осуществляется способом
У человека имплантация погружная, или интерстициальная. За одни сутки бластоциста погружается в эндометрий почти наполовину, а через 40 часов — полностью. Дефект, образовавшийся в слизистой оболочке матки, восстанавливается в течение 5 суток. Условно имплантация состоит из двух фаз — прилипания (адгезии) бластоцисты к слизистой оболочке матки и погружения (инвазии) бластоцисты в глубину слизистой оболочки.
На 6-е сутки эмбриогенеза бластоциста прикрепляется к эпителию эндометрия (обычно, эмбриональным полюсом в области задней или вентральной стенки у маточного угла). В адгезии бластоцисты важную роль играют вещества группы интегринов, вырабатываемые эпителиоцитами слизистой оболочки матки. В норме у женщин между 19-24 сутками менструального цикла — т. е. в самые оптимальные для взаимодействия с бластоцистой сроки, наблюдается активная экспрессия гена интегрина в эпителиоцитах слизистой оболочки матки.
Гаструляция
У зародыша человека гаструляция инициируется в конце 1-й недели развития, сразу по окончании дробления и сброса прозрачной зоны, когда зародыш начинает погружаться в стенку матки.
1-я фаза гаструляция продолжается всю 2-ю неделю развития. Материал внутренней клеточной массы расщепляется способом деламинации на два листка — эпибласт (верхнюю часть) и гипобласт (нижнюю часть). Клетки гипобласта — мелкие кубические, их цитоплазма имеет пенистый вид, они формируют тонкий слой под эпибластом и граничат с полостью бластоцисты. Клетки эпибласта более высокие и имеют вид псевдомногослойного призматического эпителия.
Клетки эпибласта разобщаются посредством небольших полостей, при слиянии которых в дальнейшем формируется амниотическая полость. В нижней части амниотической полости остается небольшая группа клеток эпибласта — материал будущего зародыша (эмбриобласт) и внезародышевых органов.
2-я фаза гаструляции происходит на 3-й неделе развития, осуществляется способом иммиграции и завершается формированием трех зародышевых листков — эктодермы, мезодермы, энтодермы. В будущем из материала этих листков возникнут ткани эмбриона и внезародышевых органов. Происходящие во 2-й фазе гаструляции события напоминают таковые у куриного зародыша и представителей млекопитающих.
Инициация иммиграции связана с активной и неравномерной в разных участках эпибласта пролиферацией клеток, которая приводит к формированию клеточных потоков и образованию на поверхности эпибласта структур, известных как первичная полоска и гензеновский узелок. Округлый и плоский зародышевый диск во 2-ю фазу гаструляции превращается в вытянутый с расширенным краниальным и более узким каудальным концами.
Особое морфогенетическое значение в раннем эмбриогенезе принадлежит головному отростку (нотохорду). Головной отросток растет в краниальном направлении между эпи- и гипобластом, пока не достигнет прехордальной пластинки, которая является «индикатором» будущего рта зародыша. Головной отросток дает начало развитию хорды зародыша — своеобразного клеточного стержня, который определяет первичную ось эмбриона и придает ему «жесткость».
Хорда формирует ось скелета зародыша человека и является основой развития костей осевого скелета (позвоночника, ребер, грудины, черепа). Вокруг хорды в будущем сформируется позвоночный столб. Нотохорд оказывает важное индуцирующее влияние на дифференцировку прилежащего к нему сверху участка эктодермы в нервную пластинку и далее — в нервную трубку, из которой будут развиваться головной и спинной мозг. К концу 3-й недели эмбриогенеза хорда почти полностью сформирована и простирается от орофарингеальной мембраны до каудального конца зародыша.
Источник
Гаструляция млекопитающих
Развитие млекопитающих
У высших (плацентарных) млекопитающих яйцеклетка содержит малое количество желтка и является вторичной олигоизолецитальной. Дробление полное, неравномерное, асинхронное. В ходе дробления наблюдается ранняя компактизация бластомеров, в результате чего образуется плотный клеточный шар – морула. В последующем возникает полость наполненная жидкостью – бластоцель и формируется бластоциста. В процессе образования бластоцисты бластомеры наружной клеточной массы формируют трофобласт, а бластомеры внутренней клеточной массы – эмбриобласт.
У плацентарных млекопитающих по сравнению с беспозвоночными и низшими позвоночными очень рано начинает функционировать геном зародыша. Трансляция белков с генома зародыша наблюдается со стадии восьми бластомеров.
В период дробления у млекопитающих происходит ранее обособление клеток, которые образуют внезародышевые органы, что связано с внутриутробным способом развития.

Рис. 1. Схема дробления и образования бластоцисты млекопитающих: А – полное асинхронное дробление зиготы, стадии 2 и 4 бластомеров; Б – обрастание светлыми клетками темных (морула); В – образование бластоцисты, формирование полости, обособление темных клеток внутренней клеточной массы от светлых — наружной клеточной массы — трофобласта; 1 – внутренняя клеточная масса, 2 – трофобласт
Особенности развития млекопитающих:
1) в ходе развития у млекопитающих появляется необходимость в создании таких внезародышевых органов как амнион и плацента, которые защищают зародыш и обеспечивают его метаболитами матери;
2) развитие зародыша происходит в матке, что освобождает его от необходимости развиваться быстро;
3) внезародышевые органы, в частности плацента, обеспечивают зародыша питательными веществами материнского организма, поэтому яйцеклетка не нуждается в большом количестве желтка и дробление протекает с той же скоростью что и деление соматической клетки.
Гаструляция млекопитающих, как и у всех позвоночных, протекает в две фазы и заканчивается образованием трех зародышевых листков, комплекса осевых зачатков органов и внезародышевых органов.
Первая фаза гаструляции осуществляется путем деламинации и протекает одновременно с имплантацией. При этом эмбриобласт бластоцисты расщепляется на эпи- и гипобласт (рис. 10). Эпибласт является источником развития зародыша и внезародышевой амниотической эктодермы. Клетки гипобласта перемещаются по внутренней поверхности трофобласта и участвуют в образовании желточного мешка, который плотно прилегает к трофобласту. Одновременно в конце ранней гаструляции происходит формирование таких внезародышевых органов, как желточный мешок, хорион, аллантоис.
Первыми из внезародышевых органов формируются желточный мешок и аллантоис. Сначала под эпибласт выселяются клетки, которые заполняют всю полость бластоцисты и образуют первичную внезародышевую мезенхиму. В ней формируются две полости – одна над эпибластом, вторая под гипобластом. Сформировавшиеся полости обрастают клетками за счет размножения клеток обоих клеточных пластов – эпи- и гипобласта. При этом образуются два пузыря – амниотический, расположенный над будущим зародышем, и желточный – под гипобластом; материалом для пузырей станут это будущие амнион и желточный мешок. Часть мезенхимы подходит близко к трофобласту и вместе с ним формирует хорион.
К началу ранней гаструляции зародыш имеет следующее строение: снаружи находится хорион, состоящий из двух слоев – трофобласта и внезародышевой мезенхимы. Сам трофобласт разделен на два листка – симпластотрофобласт, располагающийся снаружи, и внутренний цитотрофобласт. Полость зародыша заполнена внезародышевой мезенхимой. В ней находится два пузырька – амниотический пузырек, состоящий из внезародышевой мезенхимы и внезародышевой эктодермы; и пузырек желточного мешка, сформированный внезародышевой мезенхимой и внезародышевой энтодермой. Пузырьки прилегают друг к другу и прикреплены к хориону с помощью амниотической ножки, которая образована внезародышевой мезенхимой.
Тело зародыша, образованное клетками дна амниона и крышей желточного мешка, называется зародышевым диском и состоит из эпи- и гипобласта.
Поздняя гаструляция у млекопитающих осуществляется путем миграции и частичной инвагинации. Основные процессы протекают в эпибласте и напоминают таковые при гаструляции у птиц с образованием первичной полоски и первичного узелка. Первичная полоска содержит материал для формирования зародышевой эктодермы, и мезодермы. Первичный узелок является источником хордального отростка. Клетки первичной полоски прорывают эпибласт, мигрируют между эпибластом и гипобластом и образуют мезодерму. С этого момента зародыш становится трехслойным.
Клетки первичного узелка мигрируют под эпибласт и образуют хордальный отросток. Часть клеток первичной полоски перемещаются по направлению к гипобласту, встраиваются в него и занимают центральное положение. Из них формируется кишечная энтодерма.
Из всех зародышевых листков (но в основном из мезодермы) выселяются клетки, которые заполняют все пространство между зародышевым листком и формируют вторичную, или зародышевую, мезенхиму.
Таким образом, ранняя гаструляция у млекопитающих, как и у всех позвоночных, заканчивается расщеплением эмбриобласта на два листка, а поздняя – образованием третьего зародышевого листка – мезодермы, комплекса осевых зачатков (хорды, нервной и кишечной трубки) и внезародышевых органов (амниона, хориона, желточного мешка).
Внезародышевые органы – временные, или провизорные, – образуются у позвоночных в процессе гаструляции и имеют характерные особенности:
— формируются вне тела зародыша;
— принимают активное участие в процессе роста и развития;
— перестают функционировать при рождении.
Впервые внезародышевые органы в процессе эволюции появились у рыб в виде желточного мешка, который выполняет трофическую функцию у всех позвоночных, кроме млекопитающих. В стенке желточного мешка формируются кровеносные сосуды, которые связывают его с сосудистой системой зародыша: по ним желток, расщепленный ферментами, поступает в организм зародыша.

Рис. 2. Схема развития внезародышевых органов у птиц: 1 – скорлупа; 2 – воздушная камера; 3 – белочная оболочка (белок); 4 – амниотическая складка; 5 – зародыш; 6 – кишечная трубка; 7 – аллантоис; 8 – формирование серозной оболочки (хориона); 9 – обрастание желтка и образование желточного мешка
С выходом на сушу у позвоночных появляется необходимость еще в одном внезародышевом органе – амнионе, который дает возможность зародышу развиваться в водной среде. Необходимость водной среды для развития зародыша обусловлена тем, что она удерживает постоянную температуру окружающей среды, предохраняет зародыш от высыхания, механических повреждений, обеспечивает процессы выделения и поглощения.
Амнион (амниотическая оболочка) формируется двумя способами: при замыкании надзародышевой складки у птиц и рептилий (рис. 2) или путем раздвижения экто- и энтодермы у млекопитающих и человека (рис. 3). Независимо от способа образования зародыш оказывается окруженным амниотическими оболочками со всех сторон. Сами оболочки продуцируют жидкость, которая заполняет амниотическую полость. Химический состав амниотической жидкости изменяется по мере развития зародыша.
Функции амниотической жидкости:
— механическая защита (от повреждения, высыхания, срастания);
— выбор пути дифференцировки клеток наружной поверхности зародыша и создания необходимого давления для развития полостей – носовой, ротовой, ткани для развития легкого.
В ходе эволюции формируется орган или группа вспомогательных органов, которые обеспечивают дыхание и выделение продуктов метаболизма. Это серозная оболочка у низших и хорион, плацента и аллантоис у млекопитающих.
Серозная оболочка является самой наружной внезародышевой оболочкой, которая примыкает к скорлупе у птиц или стенке матки у млекопитающего (хорион). Эта оболочка осуществляет обмен веществ между зародышем и окружающей средой.
Аллантоис – развивается в результате выпячивания стенки задней кишки. У птиц и рептилий он выполняет дыхательную и выделительную функции. У млекопитающих проводит сосуды к зародышу и плаценте.

Рис. 3. Схема последовательных стадий образования внезародышевых органов у млекопитающих: 1 – зародыш; 2 – туловищная складка; 3 – амниотические складки; 4 – амнион; 5 – желточный мешок; 6 – аллантоис; 7 – хорион
Хорион формируется с момента исчезновения оболочки оплодотворения и внедрения трофобласта в стенку матки, то есть с момента имплантации. В том месте, где зародыш прикрепился к слизистой оболочке матки, ворсины хориона разрастаются и вместе с децидуальной оболочкой матки формируют плаценту. На остальном протяжении они редуцируют, образуя гладкий хорион.
Первоначально ворсины представляют собой выросты трофобласта и называются первичными ворсинами. Ворсины, сформировавшиеся из трофобласта и подлежащей внезародышевой мезодерме, называются вторичными. Третичные, или истинные, ворсины состоят из эпителия трофобласта, соединительной ткани и сосудов зародыша, которые прорастают вдоль аллантоиса и разветвляются в ворсины.
Сосуды ворсин связаны с сосудами зародыша через аллантоисные сосуды, которые входят в состав пупочного канатика.
Обмен метаболитами в плаценте у млекопитающих осуществляется следующим образом: метаболиты зародыша по пупочным артериям попадают в сосуды ворсин и проникают последовательно через стенку капилляра ворсины, соединительную ткань ворсины, эпителий трофобласта и поступают в материнский организм. От матери все необходимые метаболиты проходят тот же плацентарный барьер, но уже в обратном порядке и по пупочной вене поступают к зародышу.
В зависимости от типа питания зародыша у млекопитающих выделяют следующие типы плацент.
1. Эпителиохориальная – ворсины хориона врастают в ткани матки и эпителий ворсин контактирует с эпителием матки.
2. Десмохориальная – ворсины хориона разрушают эпителий маточных желез и контактируют с соединительной тканью матки.
3. Эндотелиохориальная – ворсины хориона разрушают эпителий и соединительную ткань и контактируют с эндометрием сосудов.
4. Гемохориальная – ворсины хориона разрушают стенку кровеносного сосуда матки и контактируют непосредственно с кровью материнского организма.
Животные с первым и вторым типом плацент после рождения быстро приспосабливаются к самостоятельной жизни, так как хорион только поглощает из крови матери белки и расщепляет их, а синтез собственных белков происходит в печени зародыша.
При третьем и четвертом типах плацент хорион сам синтезирует эмбриоспецифические белки из аминокислот материнского организма. Вот почему зародыши животных и человека с таким типом плацент длительное время не способны к внеутробному существованию.
Размножение млекопитающих значительно отличается от размножения других позвоночных. Огромное количество зверей живородящие. Наблюдаемая у некоторых пресмыкающихся, земноводных и даже рыб живородность существенным образом отличается от живородности млекопитающих.
Оплодотворенные яйца низших позвоночных задерживаются в яйцеводах самки, а зародыш получает все необходимые питательные вещества из запасов яйца.
В противоположность этому мелкие яйца млекопитающих имеют ничтожное количество питательных веществ. Оплодотворение у млекопитающих внутреннее. Созревшие яйцевые клетки поступают в парные яйцеводы, где происходит их оплодотворение. Оба яйцевода открываются в особый орган женской половой системы — матку. Матка — мускульный мешок, стенки которого способны сильно растягиваться. Оплодотворенная яйцеклетка прикрепляется к стенке матки, где происходит развитие плода.
В месте прикрепления яйцеклетки к стенке матки развивается плацента или детское место. Зародыш соединен с плацентой пуповиной, внутри которой проходят его кровеносные сосуды.
В плаценте через стенки сосудов из крови матери в кровь зародыша поступают питательные вещества и кислород, удаляется углекислый газ и другие, вредные для зародыша продукты жизнедеятельности. В момент рождения у высших зверей плацента отделяется от стенки матки и выталкивается наружу в виде последа.
Особенности размножения и развития отличают млекопитающих от других классов позвоночных животных.
Эволюция системы размножения зверей – от утконоса до высших млекопитающих – шла по пути совершенствования:
- органов выкармливания детёнышей молоком,
- органов развития зародыша внутри материнского организма.
Клоака, объединяющая низших зверей с пресмыкающимися и птицами, исчезает, возникает матка.
Рождение из яйца заменяется живорождением.
С точки зрения строения организма, только у высших млекопитающих есть матка и плацента – органы для обеспечения внутриутробного развития зародышей. Только млекопитающие выкармливают детёнышей молоком – веществом, которое содержит всё необходимое для развития детёныша.
Только высшие млекопитающие могут обеспечить своему потомству такую защиту, которая не повлечёт за собой сокращение числа детёнышей.
Образ жизни и поведение млекопитающих, как и других животных, меняется в связи с сезонными изменениями условий обитания. Осенью звери линяют. Неблагоприятное зимнее время многие проводят в спячке, а другие делают запасы кормов. Немногие совершают миграции. Численность млекопитающих зависит от кормовых условий, болезней, хищников.
Гаструляция у млекопитающих
Вместо того чтобы развиваться самостоятельно, большинство млекопитающих разработали замечательную стратегию развития внутри своей матери. Зародыш млекопитающих получает питательные вещества непосредственно от матери, а не пользуется накопленным желтком.
Эта эволюция повлекла за собой резкие перемены в материнской анатомии (такие, как расширение нижнего отдела яйцевода, образующее матку), а также развитие у плода органа, способного к усвоению материнской пищи — плаценты.

 Происхождение различных тканей раннего зародыша млекопитающих суммировано на рис.
Происхождение различных тканей раннего зародыша млекопитающих суммировано на рис.
4.39. Первое разделение клеток в пределах внутренней клеточной массы заключается в формировании слоя гипобласта. Эти клетки, отделяясь от внутренней клеточной массы, вселяются в полость бластоцисты, где из них образуется энтодерма желточногомешка.
Как и у зародышей птиц, у млекопитающих эти клетки не участвуют в построении какой-либо части организма новорожденного. Оставшуюся на поверхности часть внутренней клеточной массы, лежащую над гипобластом, теперь следует считать эпибластом.Эпибласт затем расщепляется посредством мелких трещин, которые в конце концов сливаются, отделяя зародышевый эпибластот других клеток эпибласта, образующих выстилку амниона. После образования выстилки амнион наполняется секретом, называемым амниотической жидкостью, которая предотвращает высыхание развивающегося зародыша.
Эмбриональный эпибласт содержит все клетки, необходимые для образования самого зародыша. В этом отношении он сходен с эпибластом птиц. На заднем крае эмбрионального эпибласта образуется локальное утолщение, в итоге дающее начало первичной полоске, через которую мигрируют клетки-предшественники энтодермы и мезодермы. Как и у зародыша птиц, клетки мигрирующие между слоями гипобласта и эпибласта, покрыты гиалуроновой кислотой.
В то время как в зародышевом эпибласте наблюдаются движения клеток, напоминающие движения клеток при гаструляции у рептилий и птиц, из клеток трофобласта образуются органы, характерные только для млекопитающихи обеспечивающие выживание плода в матке.
Часть клеток трофобласта — нормально делящиеся клетки — составляет слой, называемый цитотрофобластом,другая часть образует популяцию, в которой деления ядер не сопровождаются цитокинезом в результате чего формируется синцитиальный трофобласт.На его поверхности образуются многочисленные ворсминки.
Они внедряются в слизистую оболочку матки, закрепляя в ней зародыша. Матка в свою очередь посылает в эту область кровеносные сосуды, где они затем контактируют с синцитиальным трофобластом.
Вскоре после этого мезодермальная ткань простирается кнаружи от гаструлируюшего зародыша. К этому времени на поверхности трофобласта развиваются многочисленные выросты (ворсинки). Было показано, что мезодермальная ткань мигрирует через первичную полоску, но становится внезародышевой, а не зародышевой мезодермой и входит в ворсинки трофобласта. Из внезародышевой мезодермы формируются кровеносные сосуды, несущие питательные вещества от матери к зародышу.
Узкий соединительный стебелекиз внезародышевой мезодермы соединяет зародыш с трофобластом, а затем формирует сосуды пупочного канатика.Полностью развитый орган, состоящий из ткани трофобласта и содержащий внезародышевые кровеносные сосуды (мезодермального происхождения), называется хорионом;сливаясь со стенкой матки, хорион образует плаценту.
 На рисунке слева показана связь между зародышевыми и внезародышевыми структурами у трехнедельного плода человека.
На рисунке слева показана связь между зародышевыми и внезародышевыми структурами у трехнедельного плода человека.
Видно, что плод заключен в амнион и, кроме того, защищен еще хорионом.
Гаструляция у млекопитающих и человека
Можно легко различить кровеносные сосуды, входящие в хорион и выходящие из него, а также ворсинки, простирающиеся от наружной поверхности хориона. Эти ворсинки содержат кровеносные сосуды и увеличивают площадь хориона, доступную для материнской крови.
Для высших млекопитающих характерно более или менее плотное прикрепление зародыша к стенкам матки, наступающее через несколько дней после начала развития (у мыши на 6-е, у человека на 7-е сутки), когда зародыш находится на стадии бластоцисты.
В основе процесса прикрепления, называемого имплантацией, лежит погружение ворсинок хориона в стенку матки. В результате образуется особый орган — плацента, имеющая зародышевую часть (ворсинки хориона) и материнскую часть (более или менее измененная стенка матки).
В зависимости от расположения ворсинок на хорионе и крипт на слизистой оболочке матки у млекопитающих различают несколько типов строения плацент.
Например, у ряда копытных (коровы), лемуров и китообразных встречается полуплацента,в которой ворсинки хориона погружаются в складки слизистой оболочки матки, как пальцы в перчатку.
Такая плацента называется неотпадающей, т.к. при родах ворсинки хориона выходят из углублений слизистой оболочки матки, не повреждая ее, без кровотечения. Наиболее тесная связь сосудов плода и матери осуществляется в гемохориальной плаценте приматов и некоторых других отрядов животных. Здесь ворсинки хориона прободают эндотелий кровеносных сосудов слизистого слоя матки и непосредственно соприкасаются с материнской кровью.
Таким образом, кровь матери и плода разделена между собой лишь тонкой наружной оболочкой ворсинок хориона и стенками капиллярных сосудов зародыша.
Установлено, что клетки ворсинок хориона активно заглатывают путем пиноцитоза целые капельки крови матери. Отторжение плаценты у всех хищных, грызунов, летучих мышей и приматов при родах сопровождается отпадением части слизистой оболочки матки и кровотечением, поэтому ее называют отпадающей (децидуальной).
Таким образом, хотя кровеносные сосуды плода и матери нигде не сливаются, растворенные в крови вещества могут диффундировать через ворсинки.
Этим способом мать снабжает плод питательными веществами и кислородом, а плод отправляет в кровеносную систему матери конечные продукты обмена (главным образом диоксид углерода и мочевину).
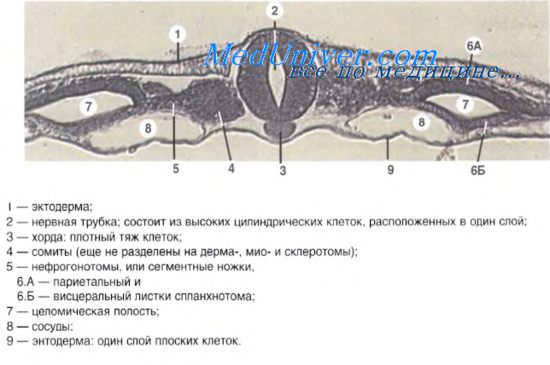
Внезародышевые органы млекопитающих и типы плацент
Формирование внезародышевых органов: оболочек, желточного мешка и аллантоиса.
Птицы, рептилии и млекопитающие относятся к группе амниот — животных, эмбриональное развитие которых протекает во внезародышевых оболочках.
Всего внезародышевых оболочек (органов) четыре:
Внезародышевые органы выполняют разные функции и различаются по тому, какие зародышевые листки участвуют в их формировании.
Амнион и хорион образуются из слоя клеток, образованно эктодермой и париетальным листком латеральной мезодермы, называемым соматоплеврой. По своему происхождению амнион и хорион являются соматоплевральными внезародышевыми оболочками.
Стенка желточного мешка и аллантоис образуются в результате разрастания слоя клеток, образованного энтодермой и висцеральным листком латеральной мезодермы – спланхноплеврой. По своему происхождению желточный мешок и аллантоис являются спланхноплевральными внезародышевыми органами.
Желточный мешок выполняет функцию органа питания.
Висцеральный листок мезодерме, входящий в состав стенки желточного мешка, развивает мощную систему кровесносных сосудов и капилляров, по которой питательные вещества из желточного мешка переносятся в тело зародыша. Жеоточный мешок соединяется со средней кишкой зародыша тонкой трубкой – желточным стебельком, внутри которого находится желточный проток.
Однако питательные вещества через желточный проток не поступают. В утилизации желтка участвуют клетки стенки желточного мешка. Сначала энтодермальные клетки расщепляют белки желтка яйца до растворимых аминокислот, которые потом поступают в кровеносные сосуды мезодермы и с кровотоком переносятся в тело зародыша.
Кроме того, желточный мешок является органом внезародышевого кроветворения. В мезодерме желточного мешка закладываются кровяные островки, которые дают первую генерацию клеток крови зародыша и являются источник стволовых кроветворных клеток. Последние мигрируют по внезародышевым кровеносным сосудам и заселяют кроветворные органы зародыша.
Амнион – внезародышевый орган,закладывается вместе с хорионом в виде складки на стенке зародышевого мешка в его головном отделе (головная амниотическая складка) в середине вторых суток инкубации.
Он состоит из внезародышевой эктодермы и париетального листка внезародышевой мезодермы. Складка из этих листков с течение третьих суток инкубации нарастает на тело зародыша до уровня поворота и вдоль его тела. На третьи сутки инкубации формируется хвостовая амниотическая складка, она начинает подниматься над хвостовой частью тела зародыша, достигая примерно уровня растущей хвостовой кишки.
К началу четвертых суток инкубации амниотические складки все больше нарастают над телом зародыша и покрывают его, образую сначала заметной величины отверстие, а затем смыкаются, оставляя узкое воронкообразное отверстие – сероамниотический проток.
Таким образом, над телом зародыша создается полость, которая через сероамниотический проток заполняется жидкой фракцией белка из белковой (третичной) оболочки яйца. С этого времени зародыш оказывается погруженным в жидкую среду, соответствующую водной среде обитания низших позвоночных животных.
Жидкость амниона примерно на 14-ые сутки инкубации становится дополнительным к желтку источником питания цыпленка непосредственно через формирующийся рот и желудочно-кишечный тракт.
Аллантоис формируется как дивертикул задней кишки зародыша, разрастающийся в полость экзоцелома и вытесняющий его. У птиц аллантоис представляет собой большой мешок, в котором накапливаются и хранятся до вылупления цыпленка токсические продукты метаболизма зародыша.
В основном это продукты азотистого обмена, накапливающиеся в виде солей мочевой кислоты.
Хорион (сероза) образует внешнюю внезародышевую оболочку эмбриона и выполняет защитную функцию. По мере разрастания аллантоиса висцеральная мезодерма, покрывающая его снаружи, прирастает к париетальной мезодерме хориона – формируется хориоаллантоис. В хориаллантоисной оболочке из материала висцеральной мезодермы формируется большое число кровеносных сосудов, формирующих густую капиллярную сеть.
Она осуществляет газообмен, то есть выполняет функцию дыхания зародыша. Хориоаллантоис обеспечивает также растворение кальция скорлупы, который необходим для остеогенеза, и его транспорт к зародышу.
 Типы плацент.
Типы плацент.
 Типы плацент.
Типы плацент. Продолжением имплантации является процесс образования плаценты – плацентация.
Плацента – провизорный орган, который формируется во время беременности, в ее построении участвуют как эмбриональные, так и материнские ткани. Через плаценту осуществляется связь зародыша с материнским организмом. Функции плаценты, обеспечивающие нормальное развитие зародыша, многообразны: трофика и газообмен, защитная, регуляторная, гормональная, антитоксическая и т.д. Основная функция плаценты заключается в передаче веществ, растворенных в крови матери, эмбриону и наоборот.
Питательные вещества диффундируют из крови матери в кровь эмбриона, а конечные продукты обмена веществ плода диффундируют в кровь матери и выводятся ее выделительной системой. Через плаценту осуществляются газообмен: кислород поступает от матери к эмбриону, а углекислый газ – от эмбриона к матери. Кроме того, плаценты многих млекопитающих вырабатывают гормоны, которые способствуют сохранению беременности (у человека это хорионический гонадотропин).
При рождении плода плацента отторгается.
У разных представителей плацентарных млекопитающих строение плаценты неодинаково. Иногда плаценты различных видов млекопитающих классифицируют по их анатомическому строению. При этом учитывается форма плаценты и расположение на ней ворсинок хориона. По этим признакам выделяют 4 вида плацент (классификация по О.Гертвигу):
7. диффузная плацента, для которой характерно образование ворсин по всей поверхности хориона;
Множественные плаценты – хориальные ворсины представлены группами, границами между ними являются участки гладкого хориона;
9. Зонарная плацента – ворсины хориона располагаются, как бы опоясывая плод;
10. Дискоидальная плацента – хориальные ворсины сконцентрированы в одном участке хориона; характерны для грызунов, приматов и человека.
Однако чаще при классификации плацент используют другой подход – гистологический.
В основу его положена степень сближения с материнским кровеносным руслом.
Кровоток плода и кровоток матери никогда не смешиваются: их разделяет несколько слоев эмбриональной и материнской ткани, которые образуют так называемый гемоплацентарный барьер.
Он обеспечивает селективный обмен растворенными веществами между системами кровообращения материнского организма и плода. Плаценты разных видов млекопитающих подразделяют на 4 типа в зависимости от количества слоев, разделяющих кровоток матери и плода:
4) Эпителиохориальные плаценты – материнские ткани не разрушаются, а ворсины хориона только прилегают к углублениям слизистой оболочки матки (свиньи, лошади и др.);
5) Синдесмохориальные плаценты – ворсины хориона разрушают эпителий матки и вторгаются в соединительную ткань слизистой оболочки матки (жвачные животные);
6) Эндотелиохориальные плаценты – ворсины контактируют с эндотелием кровеносных сосудов слизистой матки (хищники);
7) Гемохориальные плаценты – ворсины хориона соприкасаются непосредственно с материнской кровью; разветвленная сеть ворсин расположена в межворсинчатом пространстве, заполненном несвертывающейся кровью матери, то есть осуществляется наиболее совершенный контакт между плодом и материнским организмом (грызуны, насекомоядные, обезьяны и человек).
В плацентах 2-4 типов клетки трофобласта вырабатывают протеолитические ферменты, которые разрушают прилежащие маточные ткани.
Глубина проникновения клеток трофобласта в глубь материнских тканей зависит от митотической и протеолитической активности клеток трофобласта, а также мигрировать в глбь материнских тканей на ранних этапах формирования плаценты.
Клетки трофобласта вырабатывают сходные с пепсином гликопротеины, характерные для беременности (PAG – Pregnancy Associated Glycoproteins). Так, например, у жвачных в клетках трофобласта экспрессируется около 100 генов, кодирующих белки этой группы, и в результате экзоцитоза везикул, содержащих PAG.
По крайней мере часть этих молекул имеют высокую протеинкиназную активность и разрушает окружающие материнские клетки, а другая часть выполняет барьерную функцию, связывая белки, выделяемые материнскими клетками. Белки группы PAG вырабатываются и в трофобласте эпителиохориальной плаценты свиньи, но, по-видимому, они не экзоцитируются в количестве, достаточном для разршения окружающих материнских тканей.
В случае синдесмохориальных плацент (жвачные) клетки трофобласта не проходят глубоко в материнские ткани, так как довольно быстро устанавливают специализированные плотные контакты с окружающими материнскими клетками и теряют мобильность.
Клетки трофобласта, которые мигрируют в материнские ткани, являются двух- и трехъядерными, но в формировании единого слоя синцититрофобласта, как в случае гемохориальной плаценты человека, не происходит.
В случае гемохориальных плацент у приматов и человека слой синцитиотрофобласта, обладающий высокой протеолитической и фагоцитарной активностью, разрушает не только маточный эндотелий и лежащую под ним децидуальную ткань, но и стенки кровеносных сосудов, которыми пронизан эндометрий.
В гемохориальных плацентах грызунов синцитий не формируется и все клетки трофобласта сохраняют свою индивидуальность, но для них также характерны высокий уровень протеолитической и фагоцитарной активности, а на начальных этапах формировария плаценты – активная миграция в глубь материнских тканей.
В результате в обоих случаях кровь матери контактирует непосредственно с поверхностью хориона и от кровотока эмбриона ее отделяют только эмбриональные ткани.
По степени повреждения материнсаких тканей при родах все типы плацент подразделяют на отпадающие (или децидуальные) и неотпадающие.
Для свиней, лошадей, верблюдов, многих жвачных животных, лемуров и некоторых других видов плацентарных млекопитающих характерна неотпадающая плацента, состоящая только из плодных тканей.
При родах ворсины хориона выходят из углублений слизистой оболочки матки, не повреждая ее, без кровотечений.
Для хищных, грызунов, насекомоядных, рукокрылых, приматов и человека характерны плаценты отпадающего (децидуального типа). В состав такой плаценты входят как плодные, так и материнские ткани.
При родах происходит так называемое отторжение плаценты – вместе с ворсинами хориона отторгается и часть слизистой оболочки матки, что вызывает довольно значительное кровотечение.
В случаях формирования децидуальной плаценты маточный эндометрий реагирует на имплантацию эмбриона – в нем развивается децидуальная реакция. Клетки эндометрия, расположенные вокруг места имплантации, начинают активно делиться.
Источник



