
Получение гетерозисных гибридов и мужская стерильность
Несмотря на то что гетерозис наблюдается у очень многих видов растений, на практике это явление гибридной мощности используется еще у относительно небольшого числа культур. Причина этого объясняется трудностями производства гибридных семян, связанными со строением и формой репродуктивных органов у некоторых видов.
Путем обрывания початков кукурузы, являющейся однодомным растением, легко удалить мужские репродуктивные органы с материнских форм и получить таким образом достаточное количество гибридных семян. Однако кастрацию пыльников подсолнечника, риса, пшеницы, имеющих двуполые цветки, проводить трудно и экономически невыгодно. Поэтому для производства гибридных семян нужны соответствующие методы, из которых в настоящее время наиболее успешным считают метод использования мужской стерильности.
Мужская стерильность
Мужская стерильность — явление, связанное с продуцированием растениями пыльцы, не способной к оплодотворению. Она возникает вследствие полного недоразвития мужских репродуктивных органов (пыльников) или же в тех случаях, когда в нормально развитых пыльниках из-за нарушений мейоза не образуются нормальные микроспоры и пыльца становится нефункциональной, или стерильной.
Существуют три типа генетически обусловленной мужской стерильности:
1) генетическая, или ядерная, мужская стерильность, обусловленная действием одного и более генов в хромосомах;
2) цитоплазматическая мужская стерильность, обусловленная экстрахромосомными факторами, или плазмогенами;
3) цитоплазматически-ядерная мужская стерильность, обусловленная действием обоих факторов.
Из указанных типов стерильности наиболее пригодной для селекции растений оказалась цитоплазматическая мужская стерильность.
Цитоплазматическая мужская стерильность (цмс)
Этот тип стерильности спонтанно возникает у кукурузы, сахарной свеклы, моркови, лука, перца овощного и других растений. Источники цитоплазматической мужской стерильности, используемые в настоящее время, получают в основном от скрещиваний между разными видами (пшеница, рис, табак, фасоль обыкновенная, перец овощной, виды крестоцветных и др.) или даже от скрещиваний между разными родами (Aegilops ssp. X Triticum ssp.).
При скрещивании растения с цитоплазматической мужской стерильностью и растения с мужской фертильностью получают потомство с мужской стерильностью. Это показывает, что за признак цмс ответственны находящиеся в цитоплазме плазмогены. Они передаются потомству только через яйцеклетки, поскольку отцовская форма через пыльцевые зерна передает только ядро и поэтому не в состоянии вызвать у потомства мужскую стерильность. Далее установлено, что цитоплазматическая мужская стерильность как таковая не может быть самостоятельно использована в селекции гетерозисных гибридов, так как при скрещивании цмс-линий с любыми другими линиями получают стерильные гибриды F1. У лука и других растений, где от F1 используют вегетативные части, это не представляет особых трудностей, но у подсолнечника, кукурузы и других растений, основным продуктом которых являются семена, цмс использовать нельзя. Следовательно, отцовская линия (опылитель) должна содержать гены-восстановители мужской фертильности, так называемые Rf-гены, чтобы растения F1 дали семена.
Производство гибридов с использованием цмс-линий можно представить в виде следующей технологической схемы:
1) передача признака цмс той линии, которая будет служить материнской формой;
2) поддержание цмс-линии (А-линия) и ее восстановителя фертильности, или аналога (В-линия);
3) передача гена Rf линии-опылителю (R-линия);
4) производство гибридных семян путем выращивания в чередующихся рядках линий А и R.
После выделения признака цмс его передают соответствующей линии или сорту с помощью метода возвратных скрещиваний. По меньшей мере через шесть возвратных скрещиваний цмс-линии с отцовской получают A-линию, содержащую все признаки отцовской линии и дополнительно признак цмс. Поддержания A-линии добиваются ее скрещиванием с исходной линией, не имеющей цмс (В-линия), которая на самом деле является ее изогенной линией, или фертильным аналогом.
Поскольку все нормальные линии, т.е. не имеющие цмс, не несут также генов-восстановителей фертильности (Rf-гены), нужно найти такие линии, которые содержат эти гены. Обычно они находятся в части материала, относящегося к тому же виду, что и материал с цмс, но в некоторых случаях эти гены можно передавать даже путем скрещивания разных видов, как, например, перенос гена Rf из Triticum timopheevi в Tr. aestivum. Передача генов Rf линиям-опылителям (R-линии) осуществляется также с помощью возвратных скрещиваний, это необходимо у видов, используемых для получения семян. В то же время видам, у которых используют вегетативные части (лук, злаковые травы), линии-опылители с геном Rf не нужны.
Если в хромосомах линии-опылителя находится ген (или гены) Rf, это значит, что цмс-линия наряду с так называемой стерильной цитоплазмой содержит и ядерные гены, которые не в состоянии восстановить фертильность, т.е. ген (или гены) rf. Производство гибридных семян на основе цитоплазматической мужской стерильности схематически представлено на рисунке 16.1.
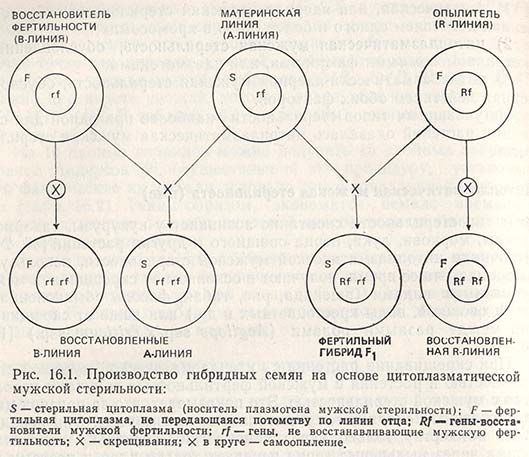
Производство гибридных семян на основе цмс таит в себе немало трудностей. Сам признак мужской стерильности проявляется недостаточно полно, так как даже в A-линиях, например, появляется некоторое количество фертильных растений. На гены rf могут сильно воздействовать факторы окружающей среды, поэтому даже растения F1 оказываются не полностью фертильными, что в конечном итоге снижает эффект гетерозиса. Наряду с изменениями, происходящими в А- и R-линиях, часто наблюдается ослабление устойчивости к патогенным организмам по сравнению с их нормальными аналогами и т.д.
Ядерная мужская стерильность
У большинства видов ядерная, или генетическая, точнее генная, мужская стерильность обусловлена одним рецессивным геном (сорго, фасоль обыкновенная), но может контролироваться и большим числом генов (ячмень, томаты). Поскольку речь идет о генах, находящихся в хромосомах, в потомствах материнской и отцовской линий происходит расщепление, и технологический процесс производства гибридных семян отличается от такового на основе цмс (рис. 16.2).
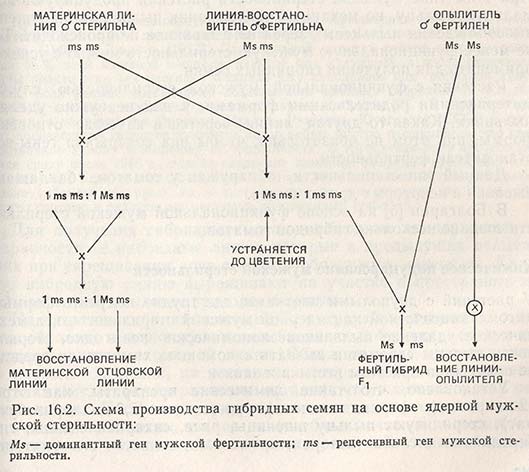
Трудности в использовании ядерной мужской стерильности состоят в следующем: во-первых, фертильные растения необходимо устранять до цветения, чтобы они не опылили мужскистерильные растения, ибо это снижает эффект гетерозиса; во-вторых, из-за доминантности гена мужской фертильности (Ms) приходится выращивать в два раза больше растений, чем это нужно для производства семян (все растения Msms устраняют).
Пытаясь преодолеть эти сложности, Рэмедж применил на ячмене так называемую сбалансированную систему третичных трисомиков, или систему ВТТ. Смысл ее заключается в получении растений-трисомиков с одной добавочной хромосомой, включающей сегмент двух негомологичных хромосом. Добавочная хромосома является носителем гена Ms, а два нормальных гомолога хромосомы содержат гены ms. Добавочная хромосома не передается потомству по отцовской линии. Потомство растения-трисомика расщепляется на
70% дисомиков (диплоидные, всегда мужскистерильные, msms) и
30% трисомиков (фертильные, Msmsms). Рэмеджу удалось передать в добавочную хромосому маркерный ген антоциановой окраски стебля, что позволило различать стерильные и фертильные растения. Однако этот метод не получил широкого применения на практике, так как он достаточно сложен; кроме того, ВТТ-растения ослаблены и дают меньше семян, чем нормальные.
Функциональная мужская стерильность
При этом типе мужской стерильности растения продуцируют нормальную пыльцу, но механизм раскрывания пыльников нарушен и высвобождения пыльцевых зерен из пыльников не происходит. Тем не менее функциональную мужскую стерильность можно с успехом применять для получения гибридных семян.
Растения с функциональной мужской стерильностью служат материнскими родительскими формами, у них не нужно удалять пыльники. Какая-то другая линия берется в качестве отцовской формы, при этом не обязательно, чтобы она содержала гены-восстановители фертильности.
Данный тип стерильности обнаружен у томатов, баклажана, ежевики и некоторых других видов. В Болгарии на основе функциональной мужской стерильности создано несколько гибридов томатов.
Химическое индуцирование мужской стерильности
У растений с двуполыми цветками, где трудно получить формы с цитоплазматической или ядерной мужской стерильностью, а механическое удаление пыльников экономически невыгодно, стерильность пыльцы стремятся вызвать с помощью химических средств, не повреждая при этом пестика и завязи.
Установлено, что такие химические препараты, как этрел (2-хлорэтанфосфорная кислота) или мендок (2,3-дихлоризобутират), стерилизуют пыльцу пшеницы, риса, сахарной свеклы; гибберелловая кислота (которая, кроме того, является стимулятором роста) обладает таким же действием на салат, лук, подсолнечник и т.д. Все эти вещества называют общим термином гаметоциды.
Успешное индуцирование мужской стерильности с помощью гаметоцидов позволило бы удешевить и ускорить товарное производство гибридных семян без применения длительной процедуры передачи плазмогена цмс и гена Rf. Однако современные гаметоциды имеют ряд отрицательных эффектов. Прежде всего они вызывают неполную стерильность пыльцевых зерен; к тому же их активность зависит от доз и сроков обработок, которые необходимо повторять несколько раз за вегетацию. Гаметоциды также вызывают повреждения женских генеративных органов, приводят к деформации самих растений и удлинению вегетационного периода. По всем этим причинам они не получили еще широкого признания в практике, но тем не менее работа по созданию новых гаметоцидов и по усовершенствованию методов их применения продолжается.
Источник
Способ получения гибрида растения подсолнечника




Владельцы патента RU 2302105:
Изобретение относится к области сельского хозяйства, в частности к способу получения гибридов подсолнечника. Получают самостерильные растения из самофертильных путем изменения условия их выращивания. Далее проводят скрещивание полученных самостерильных растений с растениями, обладающими мужской фертильностью или выращенными в не изменявшихся условиях. Для получения самостерильных растений берут самофертильные растения и выращивают их в обычных для них условиях до фазы цветения. С началом цветения самофертильных растений для них устанавливают 24-часовой световой день на весь период цветения корзинки (обычно это 5-7 дней). Полученные таким образом самостерильные растения используют в качестве материнской формы для скрещивания с мужски фертильными. Изобретение позволяет повысить эффективность получения материнских растений. 2 з.п. ф-лы, 3 табл.
Изобретение относится к сельскому хозяйству, а именно к селекции и генетике возделываемых растений подсолнечника.
Для надежной гибридизации растений требуется предварительное получение материнских, т.е. одновременно женски фертильных и мужски стерильных, организмов, дающих гибридные семена только при опылении пыльцой отцовского растения и не обладающих способностью к самоопылению. При этом следует различать мужскую стерильность, как неспособность образовывать жизнеспособные мужские гаметы, и самостерильность, как неспособность к самооплодотворению при нормальности мужских и женских гамет.
Известен способ получения гибридов у культурного подсолнечника Helianthus annuus L., являющегося однодомным растением с двуполыми протерандрическими трубчатыми цветками, собранными в соцветие-корзинку, на основе ручной кастрации, согласно которому для получения материнских растений пыльники каждого цветка удаляют пинцетом в первый день его цветения с 6 до 8 часов утра с последующим нанесением пыльцы отцовского растения на рыльца пестиков кастрированных цветков (Гундаев А.И. Основные принципы селекции подсолнечника. В кн.: Генетические основы селекции растений. — М.: Наука, 1977. — С.417-465).
Недостатком известного способа является большая трудоемкость, необходимость квалифицированного труда, ненадежность процесса гибридизации из-за возможного самоопыления при неполном удалении или растрескивании пыльников.
Известен способ получения мужски стерильных аналогов растений подсолнечника на основе использования экзогенного воздействия экологических факторов, а именно химической кастрации, согласно которому растения индивидуально опрыскивают водным раствором гиббереллина в фазе розетки во время закладки генеративных органов корзинки при шести-восьми парах листьев. Урожайность снижается при этом до уровня 70% (а.с. СССР №211926, А01Н 3/04, 1968, №8).
Недостатком известного способа является большая трудоемкость и снижение женской фертильности обработанных гаметоцидом растений.
Известен также способ получения гибридов растений, взятый нами за прототип (Патент РФ №2210884, А01Н 1/02, публ. 2003, бюл. №24), согласно которому изменяют условия выращивания растений, чтобы сделать их мужски стерильными, затем скрещивают полученные мужски стерильные растения с растениями, обладающими мужской фертильностью или выращенными в не изменявшихся условиях, для получения гибридов, при этом изменение условий роста растений заключается в изменении одного или нескольких факторов их выращивания, таких как место выращивания, период выращивания, влажность, питание, фотопериод и температурные условия, причем изменение фотопериода заключается в выращивании растений короткого дня в условиях с фотопериодом, составляющим 12 ч/день и более, либо в выращивании растений длинного дня в условиях с фотопериодом менее 12 ч/день.
Недостатком известного способа является низкая эффективность выхода мужски стерильных растений на уровне 15% за счет изменения одного определенного условия выращивания.
Настоящим изобретением решается задача увеличения эффективности получения материнских растений.
Цель изобретения — получение мужской самостерильности трубчатых цветков корзинки подсолнечника за счет достижения морфологической женской однополости этих цветков, обеспечивающей им неспособность к самоопылению.
Технический результат достигается тем, что в известном способе получения гибрида растения, включающем скрещивание растения, мужскую стерильность которого получают изменением условий выращивания, с растением, обладающим мужской фертильностью или выращенным в не изменяющихся условиях, причем в качестве фактора для изменения условия выращивания растений используют фотопериод, согласно изобретению для получения мужской стерильности растение выращивают с фотопериодом 24 часа светового дня в фазу цветения корзинки, причем до начала цветения растения выращивают в обычных условиях, а 24-часовой световой день устанавливают на весь период цветения корзинки — от 5 до 7 дней.
Сопоставительный анализ заявляемого технического решения с прототипом позволяет сделать вывод, что заявляемый способ отличается от известного условием осуществления способа — режимом его осуществления: в известном способе изменение фотопериода для растений короткого дня осуществляют в сторону увеличения от 12 ч/день, а для растений длинного дня — в сторону уменьшения от 12 ч/день, причем такой режим устанавливают на весь период выращивания, а в заявляемом способе устанавливают 24-часовой световой день и только в фазу цветения корзинки подсолнечника.
Таким образом, заявляемый способ соответствует критерию патентоспособности «новизна».
Исследуя уровень науки и техники в процессе проведения патентного поиска по всем видам сведений, общедоступных в печати, мы не обнаружили решений, имеющих признаки, совпадающие с отличительными. Опубликованных данных о получении мужски стерильных растений подсолнечника при фотопериодическом воздействии в фазе цветения корзинки не обнаружено. Кроме того, отличительные от прототипа признаки, содержащиеся в независимом пункте формулы изобретения, для специалиста в области селекции и генетики явным образом не следуют из известного на сегодня существующего уровня техники.
Все известные способы получения мужски стерильных растений предполагают воздействие в период роста растений, а именно в начале формирования генеративных органов, в частности в фазе розетки, т.е. на ранних критических стадиях развития пыльников. Более того, выращивание подсолнечника при непрерывном освещении в условиях камер искусственного климата не влияет на переход растений к цветению, незначительно сокращает вегетационный период на 3-7 дней (Клюка В.И. и Цуркани С.Н. Интенсивность и продолжительность освещения при выращивании подсолнечника в условиях искусственного климата. В кн.: Вопросы физиологии масличных растений в связи с задачами селекции и агротехники.» Краснодар: Изд-во ВНИИМК, 1975. — С.101-107). Наши исследования доказали возможность получения мужски самостерильных женски фертильных растений за счет воздействия только на процесс цветения корзинки 24-часовым днем, которые могут использоваться в получении гибридов в качестве материнской формы при отсутствии самоопыления.
Поэтому можно сделать вывод о соответствии заявленного изобретения критерию патентоспособности «изобретательский уровень».
Заявляемое техническое решение соответствует и критерию патентоспособности «промышленная применимость», так как оно может быть использовано в селекционной практике для создания экспериментальных гибридов подсолнечника в камерах и теплицах фитотрона и, кроме того, в описании изобретения показаны средства и методы, с помощью которых возможно осуществление технического решения в том виде, как оно охарактеризовано в независимом пункте формулы изобретения.
Способ осуществляют следующим образом.
Для получения мужски стерильного растения подсолнечника растение самофертильной линии выращивают в камерах или теплицах фитотрона до стадии цветения в обычных условиях (с фотопериодом 16 часов день/8 часов ночь). С началом цветения растения устанавливают 24-часовой световой день на все время цветения корзинки (от 5 до 7 дней). Получаемое мужски стерильное растение с морфологически однополыми женскими цветками за счет невыхода пыльников из венчика используют в качестве материнской формы и скрещивают с растением, обладающим мужской фертильностью или выращенным в не изменявшихся условиях.
Пример. Растения константной селекционной линии подсолнечника с генетически измененным составом жирных кислот и токоферолов ВК876 выращивали летом 2004 г. в 16 сосудах с почвой в теплице при естественном освещении стандартным методом. После начала цветения 12 растений были перенесены в камеру фитотрона с 24-часовым световым днем при использовании светильников типа Фотос 4 с лампами ДРИ-2000-6 и освещенности 25 килолюкс. Четыре растения оставлены в теплице в качестве контроля. Для всех 12 растений в камере фитотрона наблюдали нарушение процесса цветения трубчатых цветков в виде отсутствия явления протерандрии для концентрической зоны соответствующего дня цветения, заключающегося в невыходе пыльниковой трубки с пыльцой из венчика за счет полного отсутствия роста тычиночных нитей (таблица 1). Длина венчика и длина пыльниковой трубки достоверно не различались. При этом наблюдали обычный рост пестика с последующим раскрытием двух лопастей рыльца без выноса пыльцы.
Искусственное вскрытие венчиков этих морфологически женских однополых цветков в трех корзинках показало, что нормально развитые пыльниковые трубки содержали обычное количество пыльцы. В случае возвращения цветущего в течение 2-3 дней растения в условия обычного фотопериода цветки оставшейся части корзинки показали, также, как и оставленные контрольные растения, нормальное протерандрическое развитие с выталкиванием пыльцы из предварительно проросшей пыльниковой трубки сомкнутыми лопастями рыльца пестика. Следовательно, фотопериодически детерминированная морфологическая женская однополость проявляется только у цветков зоны цветения, находившейся под воздействием 24-часового светового дня.
Три растения линии ВК876 с фотопериодической морфологической женской однополостью цветков в камере фитотрона индивидуально опыляли пыльцой линии ЛГ28, другие три растения опыляли пыльцой линии ВК876, полученной при обычном фотопериоде, оставшиеся три растения изолировали для самоопыления. Завязываемость семян при опылении шести корзинок варьировала от 90 до 98%, что свидетельствует о высокой женской фертильности материнских растений. Все три растения, оставленные для самоопыления, не образовали семян, т.е. оказались полностью самостерильными.
Изоляция отдельных корзинок самофертильной линии ВК876 в условиях нормального фотопериода теплицы привела к формированию семян от самоопыления (таблица 2). Относительно меньшее число гибридных семян на корзинку по отношению к числу семян, полученных от самоопыления, объясняется одноразовым нанесением пыльцы отцовской формы на рыльца пестиков в скрещивании, тогда как самоопыление продолжалось все пять дней цветения корзинки.
Дополнительным доказательством гибридности, а не самоопыленности, полученных семян в скрещивании ВК876 ФМС × ЛГ28 явилось изменение биохимических маркеров семян. Так содержание олеиновой кислоты достоверно снизилось с 84 до 80% от суммы жирных кислот масла, а содержание α-формы в токоферольном комплексе увеличилось до 100% (таблица 3).
Таким образом, в предлагаемом способе выход самостерильных растений после фотопериодического воздействия на самофертильные растения составил 100%, степень гибридизации — 100% при полном отсутствии самоопыления материнских растений.



Итак, мужская самостерильность трубчатых цветков достигается фотопериодическим воздействием 24-часового светового дня в фазу цветения корзинки, вызывающим нарушение процесса цветения андроцея в виде отсутствия протерандрии для концентрической зоны соответствующего дня цветения, заключающегося в невыходе пыльниковой трубки с пыльцой из венчика цветка за счет отсутствия роста тычиночных нитей при нормальном росте пестика с последующим раскрытием двух лопастей его рыльца без пыльцы.
В случае необходимости получения пыльцы с материнского растения для его самоопыления или использования в качестве отцовской формы следует перейти на 16-часовой фотопериод, что приведет к обычному типу цветения оставшейся части корзинки с выходом пыльников из вновь распустившихся цветков и выталкиванием пыльцы из пыльниковой трубки лопастями рыльца пестика. При этом не вышедшие из венчика пыльники предыдущих концентрических зон цветения уже не прорастают.
Следовательно, указанная совокупность заявленных признаков является необходимой и достаточной для достижения поставленной цели (получение мужской самостерильности трубчатых цветков корзинки подсолнечника за счет достижения морфологической женской однополости этих цветков, обеспечивающей им неспособность к самоопылению) и решения поставленной задачи (увеличение эффективности получения материнских растений).
1. Способ получения гибрида растения подсолнечника, включающий скрещивание растения, мужскую стерильность которого получают изменением условий выращивания, с растением, обладающим мужской фертильностью или выращенным в не изменявшихся условиях, причем в качестве фактора для изменения условия выращивания растений используют фотопериод, отличающийся тем, что для получения мужской стерильности растение выращивают с фотопериодом 24 ч светового дня в фазу цветения корзинки.
2. Способ по п.1, отличающийся тем, что до начала цветения растение выращивают в обычных условиях.
3. Способ по п.1, отличающийся тем, что 24-часовой световой день устанавливают на весь период цветения корзинки — от 5 до 7 дней.
Источник



