
База знаний
Амебоидное движение
Амебоидное движение является широко распространенной формой клеточного движения. Им обладают самые разнообразные клетки — простейшие из класса саркодовых, зооспоры, некоторые сперматозоиды (у аскариды) и яйцеклетки, плазмодии миксомицетов, фибробласты и лейкоциты, эпителиальные и нервные клетки в тканевых культурах, клетки эмбрионов позвоночных. Хорошо выражено амебоидное движение у миобластов, из которых и развиваются мышечные волокна. При регенерации эпителия клетки становятся подвижными и путем амебоидных движений перемещаются в глубь раны. Одним из способов распространения злокачественных новообразований внутри организма является амебоидное движение раковых клеток.
Амебоидное движение состоит в медленном перетекании тела клетки по субстрату и осуществляется благодаря внутриклеточному течению цитоплазмы и образованию временных псевдоподий (ложноножек). В наиболее простом случае образуется всего лишь одна Псевдоподия. Поверхностный слой клетки — эктоплазма (кортикальный или гиалиновый слой) — обладает высокой вязкостью и является, по-видимому, гелем. При образовании псевдоподий происходит превращение вязкого гиалинового слоя в легкотекучий золь, а более жидкая центральная часть протоплазмы (эндоплазма), существующая в форме белкового золя, перетекает в направлении движения в образующуюся псевдоподию. Эндоплазма в плазмодии слизевика образует одновременно несколько потоков, движущихся в разных направлениях.
Скорости амебоидного движения некоторых клеток представлены в таблице 17.
| Таблица 17. Скорость амебоидного движения | |
| Клетки | Скорость, мкм/с |
| Свободно ползающие амебы | 0,5—4,5 |
| Амебоидные зародыши слизистых споровиков | 0,33—1,3 |
| Нейтрофильные лейкоциты | 0,58 |
| Неполяризованные макрофаги | 0,004 |
Скорость движения зависит от температуры и кислотности среды, от осмотического давления, от соотношения концентрации одновалентных и двухвалентных катионов. Недостаток кислорода не прекращает амебоидного движения, но замедляет его. Под влиянием любого сильного раздражителя (0,1 M KCl, нагревание до 40°C, встряхивание) амеба сокращается, тело ее округляется, она теряет способность перемещаться по субстрату и изменять форму тела, а затем прекращается и движение гранул цитоплазмы.
Большинство современных теорий амебоидного движения являются контрактильными, объясняющими возникновение движущей силы за счет сокращения контрактильных структур цитоплазмы, в качестве которых рассматриваются микротрубочки и микронити, обнаруженные у многих видов амеб. Эти структуры могут либо свободно находиться в цитоплазме, либо образовывать агрегаты толщиной до 40 нм и более. Расположены они и в плазмалемме, и в эктоплазме на границе с эндоплазмой в виде параллельных пучков или в виде сетки, распространяющейся и в эндоплазму. Биохимическими исследованиями показано, что цитоплазматические фибриллы амеб содержат сократительные белки, подобные миксомиозину. Именно благодаря им глицеринизированные амебы и фибробласты реагируют на действие АТФ длительным сокращением всего тела.
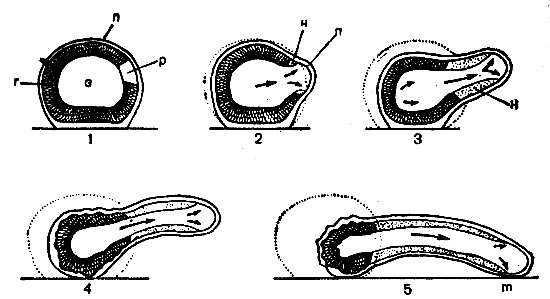
Рис. 41. Схема образования псевдоподии у амебы согласно контрактильной гипотезе (Jdhn, Bovee, 1969): 1 — превращение геля (г) в золь (з) в области p; 2 — сокращение геля развивает силу (стрелки), вытесняющую золь по направлению к плазмалемме (п), Н — новые места образования геля; 3, 4, 5 — дальнейшие стадии удлинения псевдоподии, прикрепляющейся в m
Авторы разных теорий амебоидного движения не согласны лишь относительно места, где возникают эти движущие силы. Одни считают, что амеба передвигается, «выдавливая» себя в псевдоподию за счет давления, создаваемого сокращением кортикального геля в заднем конце клетки, которое и заставляет течь жидкую эндоплазму (рис. 41). Эта гипотеза основывается на сократительных свойствах эктоплазмы, состоящей из актомиозинового геля, который, уплотняясь, сокращается.
Согласно представлению других, цитоплазма подтягивается вперед в образующуюся псевдоподию благодаря сокращению эндоплазмы в переднем конце клетки. Некоторые исследователи полагают, что силы, необходимые для движения протоплазмы, локализованы в самих псевдоподиях и возникают благодаря активному скольжению гелевых нитей или эндоплазмы по поверхности кортикального слоя. Подчеркивается также роль активного растяжения или расслабления плазмалеммы в амебоидном движении.
Амебоидное движение следует рассматривать как сложный многоступенчатый процесс, включающий и течение цитоплазмы (циклоз), и изменение ее состояния (золь ⇔ гель), а также плазмалеммы и формы тела, и образование новых псевдоподий, и, наконец, прикрепление к субстрату. Поэтому механизм амебоидного движения нельзя свести только к механизму течения цитоплазмы, он, несомненно, является гораздо более сложным (Серавин, 1967).
Источник
40. Типы органоидов передвижения простейших. Строение органоидов передвижения простейших.
Различают пять основных форм движения простейших: амебоидное, жгутиковое, ресничное, метаболирующее и скользящее.
Оно выражается в образовании ложных ножек (псевдоподий) — выростов цитоплазмы разной формы. Все содержимое клетки как бы медленно перетекает в направлении образующейся псевдоподии, и таким путем осуществляется перемещение простейшего в пространстве.
Жгутик/ресничка включает свободную часть или ундулиподию, содержащую аксонему, переходную зону, кинетосому и корешки.
Ундулиподия снаружи одета мембраной, являющейся продолжением клеточной мембраны. Поверхность ундулиподии может быть гладкой, в других случаях ундулиподия на своей поверхности несет ультрамикроскопические выросты разного строения (чешуйки или палочковиджные выросты — мастигонемы). Внутри ундулиподии расположена аксонема. У части видов параллельно аксонеме по всей длине жгутика проходит параксиальный тяж (эвгленовые, кинетопластиды). Параксиальный тяж состоит из микрофиламентов и, вероятно, увеличивает упругость жгутика.
Аксонема представляет собой цилиндр, стенка которого построена из 9 пар микротрубочек (периферические микротрубочки). В центре этого цилиндра располагаются две центральные микротрубочки (структура 9+2). Периферические микротрубочки, составляющие пару (дублет), неодинаковы, их называюют А и В. От микротрубочек А отходят выросты (динеиновые ручки), направленные к соседнему дублету, а также радиальные спицы — к центральной паре микротрубочек.
Переходная зона расположена на уровне выхода жгутика из клетки, в ней изменяется структура аксонемы (дублеты сменяются триплетами) и появляются дополнительные фибриллярные образования. Часто в этой зоне присутствуют разнообразные опорные структуры.
Кинетосома лежит под основанием жгутика/реснички, она представляет собой полый цилиндр, состоящий из 9 триплетов микротрубочек. Большинство жгутиконосцев имеет 2 жгутика и, соответственно, 2 кинетосомы. У одножгутиковых видов кинетосом тоже 2, но одна из них непосредственной связи со жгутиком не имеет. Взаимное расположение двух кинетосом различно в разных группах.
С кинетосомами связаны корешки, которые могут быть фибриллярными или микротрубочковыми, характеризуются разным размером и расположением. В целом корешки выполняют цитоскелетную функцию.
Движущим началом работы жгутика/реснички являются периферические микротрубочки и их ручки, которые обладают АТФ-азной активностью. За счет выделяющейся энергии динеиновые ручки одного дублета прикрепляются к соседнему дублету и сгибаются. В результате этот дублет смещается по отношению к другому. Последовательное повторение цикла обеспечивает поочередное смещение дублетов что и вызывает изгибание жгутика. Центральные фибриллы играют опорную роль.

Механизм жгутикового движения у разных видов различен. В большинстве случаев он сводится к вращательному движению. Жгутик описывает фигуру конуса, вершиной обращенного к месту его прикрепления. Нередко вращательное движение жгутика сочетается с его волнообразным движением. Обычно при поступательном движении само тело простейшего вращается вокруг продольной оси. У многожгутиковых движение жгутиков может носить иной характер, в частности жгутики могут находиться в одной плоскости, не образуя конуса вращения.
Этот тип движения характерен для эвгленовых и осуществляется путем активного скольжения белковых пластин покровов относительно друг друга. Внешне такое движение выглядит как волнообразное изгибание тела клетки.
Скольжение представляет собой однонаправленное движение без изменения формы клетки, этот тип движения характерен для грегарин. На поверхности тела грегарин расположены продольные складки (эпицитарные гребни), в вершинах которых проходят микрофиламенты. Волнообразные (ундулирующие) движения этих эпицитарных гребней обеспечивает скользящее движение клетки. В свою очередь движения эпицитарных гребней, вероятно, обусловлены взаимодействиями микрофиламентов с внутренним мембранным комплексом пелликулы, что приводит к сдвигу этого мембранного комплекса относительно плазмолеммы и временной деформации гребня.
Источник
Зоология беcпозвоночных
ПОДЦАРСТВО ПРОСТЕЙШИЕ,
ИЛИ ОДНОКЛЕТОЧНЫЕ (PROTOZOA)
К подцарству одноклеточных относят животных, тело которых состоит из одной клетки. Морфологически они сходны с клетками многоклеточных животных, но физиологически отличаются тем, что кроме обычных функций клетки (обмен веществ, синтез белка и др.) они выполняют функции целостного организма (питание, движение, размножение, защита от неблагоприятных условий среды). Отдельные функции у многоклеточных организмов выполняются специальными органами, тканями или клетками, а у одноклеточных функции организма выполняют структурные элементы одной клетки — органеллы. Деление клеток у многоклеточных животных приводит к росту организма, а у простейших — к размножению.
Таким образом, простейшие — это организмы на одноклеточном уровне организации. Целостность организма простейших поддерживается функциями одной клетки, а у многоклеточных — за счет взаимодействия клеток, тканей и органов.
Жизненный цикл простейших складывается из фаз развития с одноклеточной организацией, а у многоклеточных чередуются одноклеточные фазы развития с многоклеточными.
В настоящее время известно более 39 тыс. видов простейших, однако ежегодно обнаруживаются десятки и сотни новых видов, что является показателем недостаточной изученности этой группы животных.
Впервые простейшие были обнаружены голландским ученым А. ван Левенгуком — первым изобретателем микроскопа (1675). Его микроскопы представляли собой сильно увеличивающие лупы, которые давали увеличение в 100 и даже в 200 раз. Особенно много простейших первые микроскописты обнаруживали в настоях трав (infusum — означает «настойка»), поэтому первое время этих животных называли «настоечными» или инфузориями. Теперь это название сохранилось лишь за одной группой простейших. В первой системе животных К. Линнея (1759) простейшие были отнесены к одному роду — Chaos — класса червей. Только
в XIX в. Келликер и Зибольд их выделили в самостоятельный тип (1845). На Международном конгрессе протозоологов в 1977 г. была принята новая система простейших, отразившая последние достижения науки. Согласно новым принципам, опубликованным в 1980 г. (Левайн и др.), простейшие объединены в подцарство одноклеточных и подразделены на семь типов.
Общая характеристика простейших. Простейшие широко распространены в различных средах. Большинство простейших — обитатели морей и пресных вод. Некоторые виды обитают во влажной почве. Множество простейших паразитируют в других организмах. Экологическая радиация простейших отражена на рисунке 15.
Большинство простейших — мелкие организмы. Их средние размеры измеряются несколькими десятками микрометров (1 мкм равен 0,001 мм). Самые мелкие простейшие — внутриклеточные паразиты достигают всего 2- 4 мкм, а длина самых крупных видов, например некоторых грегарин, может достигать 1000 мкм. Ископаемые раковинные корненожки, например нуммулиты, в диаметре достигали 5- 6 см и более.
Форма тела простейших чрезвычайно разнообразна. Среди них имеются виды с непостоянной формой тела, как амебы. Разнообразны типы симметрии у простейших. Широко распространены формы с радиальной симметрией: радиолярии, солнечники. Это в основном плавающие планктонные простейшие. Двусторонняя симметрия наблюдается у некоторых
жгутиковых, фораминифер, радиолярий. Поступательно-вращательная симметрия характерна для фораминифер со спиральнозакрученной раковиной. У некоторых видов наблюдается метамерия — повторяемость структур по продольной оси. Разнообразны жизненные формы простейших, или морфоадаптивные типы. Наиболее широко распространенными формами являются: амебоидных, которые ведут ползающий образ жизни на различных субстратах в воде или в жидкой среде в теле хозяина; раковинные — малоподвижные бентосные формы; активно плавающие жгутиконосцы и ресничные, парящие в составе планктона радиальные, или лучистые, формы; сидячие — стебельчатые, узкотелые или плоскотелые скважники субстратов — интерстициалы, а также округлые неподвижные, покоящиеся формы (цисты, споры).
Строение клетки простейших характеризуется всеми основными признаками клеточного строения эукариот. Ультраструктура строения простейших изучена биологами благодаря использованию электронно-микроскопической техники. Разрешающие способности современного электронного микроскопа позволяют получать увеличение в 200- 300 тыс. раз.
Клетка простейших типична для эукариотных организмов и состоит из цитоплазмы и одного или нескольких ядер. Цитоплазма ограничена снаружи трехслойной мембраной. Общая толщина мембраны около 7,5 наномикрон (1 нм = 10 — 6 мм). В цитоплазме простейших различают наружный, более прозрачный и плотный слой — эктоплазму и внутренний, зернистый слой — эндоплазму. В эндоплазме сосредоточены все основные органеллы клетки: ядро, митохондрии, рибосомы, лизосомы, эндоплазматическая сеть, аппарат Гольджи и др. Кроме того, у простейших имеются особые органеллы: опорные, сократительные фибриллы, пищеварительные и сократительные вакуоли и др. Ядро покрыто двуслойной мембраной с порами. Внутри ядра находится кариоплазма, в которой распределены хроматин и ядрышки. Хроматин представляет собой деспирализованные хромосомы, состоящие из ДНК и белков типа гисто-нов. Ядрышки подобны рибосомам и состоят из РНК и белков. Ядра простейших разнообразны по составу, форме, размерам.
У простейших можно выделить особые функциональные комплексы органелл, которые соответствуют системам органов и тканей многоклеточных.
Покровные и опорные органеллы. Часть видов одноклеточных не обладает покровными и опорными структурами. Клетка таких простейших ограничена лишь мягкой цитоплазматической мембраной. Такие виды не имеют постоянной формы тела (амебы). У других видов имеется плотная эластичная оболочка — пелликула, образующаяся за счет уплотнения периферического слоя эктоплазмы и наличия в нем различных
опорных фибрилл. В этом случае простейшие обладают определенной формой тела (инфузории, эвглены) и вместе с тем они сохраняют гибкость и могут изгибаться при движении, частично сокращаться. Другие одноклеточные выделяют снаружи панцирь из чешуек, что препятствует изменению формы тела (диатомовые жгутиковые). Форму тела дополнительно могут поддерживать и другие опорные структуры — фибриллы, образующие, например у некоторых инфузорий, кортекс.
К опорным образованиям относится еще и скелет. Скелет простейших может быть наружным (раковина) или внутренним (скелетные капсулы, иглы). Раковина выделяется эктоплазмой клетки, и при этом образуется внеклеточное образование, имеющее защитную функцию. Внутренний скелет образуется в эндоплазме клетки. Формирование скелетных капсул и игл происходит путем биокристаллизации. Скелетные образования состоят из органических и минеральных веществ. Чаще всего скелеты простейших включают карбонат кальция (СаСО3) или оксид кремния (SiO2), реже сульфат стронция (SrSO4).
Двигательные органеллы. Наиболее примитивным способом движения у простейших можно считать амебоидное движение при помощи ложных ножек, или псевдоподий. При этом образуются особые выступы клетки, в которые перетекает цитоплазма. Такие органеллы движения присущи одноклеточным с непостоянной формой тела.
Более сложное движение свойственно простейшим, имеющим в качестве органелл движения жгутики или реснички. Строение жгутика и ресничек сходно (рис. 16). Каждый жгутик снаружи покрыт трехслойной цитоплазматической мембраной. Внутри жгутика имеются фибриллы: две центральные и девять двойных периферических. Жгутик крепится в цитоплазме при помощи базального тельца — кинетосомы. Обычно жгутики производят вращающее движение, а реснички — гребное. Жгутики свойственны жгутиконосцам, а реснички — инфузориям.
Некоторые простейшие способны к быстрому сокращению тела за счет особых сократительных фибрилл — мионем. Например, сидячие инфузории — сувойки способны резко сокращать свой длинный стебелек и сворачивать его в спираль. Радиолярии способны то растягивать тело клетки на радиальных иглах, то сокращать его за счет сократительных волокон. Это обеспечивает им регуляцию свободного плавания в толще воды. При неблагоприятных условиях многие простейшие инцистируются, т.е. выделяют вокруг себя плотную оболочку и превращаются в цисту.
Среди простейших немало внутриклеточных паразитов, ведущих неподвижный образ жизни и не имеющих органелл движения.
Типы питания и трофические органеллы. По типу питания простейшие разнообразны. Среди них имеются автотрофы, способные к фотосинтезу. Это одноклеточные водоросли из жгутиковых. У них имеются в цитоплазме хлорофилловые зерна, или хроматофоры.
Большинство простейших гетеротрофы, питающиеся как животные, готовыми органическими веществами. Часть из них обладает голозойным способом питания, проглатывая твердые комочки пищи. Другие питаются сапрофитным способом, поглощая растворенные органические вещества. Частицы пищи заглатывают амебы, инфузории. У них в цитоплазме образуются пищеварительные вакуоли, где происходит переваривание пищи. Такое заглатывание твердой пищи клеткой получило название фагоцитоза. При сапрофитном способе питания пищеварительные вакуоли не образуются. Однако известно, что многие простейшие могут заглатывать жидкость через временное впячивание мембраны — особую воронку. Такое поглощение жидкости называется пиноцитозом.
Некоторые виды обладают смешанным типом питания (миксотрофы). Они способны к фотосинтезу, как растения, и к питанию готовым органическим веществом, как животные. У них имеются в цитоплазме хлорофилловые зерна, но могут образовываться и пищеварительные вакуоли. К таким простейшим со смешанным типом питания относятся, например, эвглены, питающиеся на свету как растения, а в темноте как животные.
Органеллы выделения и осморегуляции. Выделение и осморегуляция осуществляются у простейших сократительными вакуолями. Они имеются только у пресноводных форм и отсутствуют у морских и паразитических видов, живущих в изотонической среде. Сократительная вакуоль в простейшем случае представляет собой пузырек в цитоплазме, регулярно заполняющийся жидкостью, которая затем удаляется наружу через пору в мембране клетки. Постоянное удаление избытка воды из клетки позволяет регулировать осмотическое давление в цитоплазме. Выделение продуктов обмена происходит у большинства простейших через поверхность клетки, а также через сократительную вакуоль, если она имеется. Особых органелл дыхания у них нет, и они поглощают кислород через клеточную мембрану.
Ядерный аппарат состоит из одного или нескольких ядер. Ядра регулируют обменные процессы клеток простейших и обеспечивают размножение. Ядра простейших варьируют по форме, числу, плоидности, функциям. У некоторых многоядерных простейших различают два типа ядер: генеративные и вегетативные. Это явление получило название ядерного дуализма. Вегетативные ядра регулируют все жизненные процессы в клетке, а генеративные участвуют в половом процессе. Ядерный дуализм характерен для инфузорий, некоторых фораминифер. Ядра простейших могут быть гаплоидными на определенном этапе жизненного цикла, или диплоидными, или полиплоидными. Большинство простейших одноядерные (моноэнергидные). Виды, у которых много ядер, называют полиэнергидными.
При бесполом размножении простейших ядра делятся путем митоза. Ядра простейших, для которых известен половой процесс, претерпевают мейоз, или редукционное деление. В отличие от многоклеточных, мейоз у одноклеточных разнообразен. В примитивном случае мейоз осуществляется в процессе одного деления клетки, в других, как у высших животных, в результате двух последовательных делений. В одних случаях редукционное деление происходит после образования зиготы (зиготическая редукция), в других, как у многоклеточных, при формировании гамет (гаметическая редукция).
Типы размножения простейших разнообразны. Им свойственно бесполое и половое размножение. Бесполое размножение осуществляется путем деления клетки на две или множество клеток (агамогамия) при митотическом делении ядер. Половое размножение простейших характеризуется образованием половых клеток — гамет (гамогамия) с их последующим слиянием (копуляция), что приводит к формированию зиготы, из которой развивается новый дочерний организм. У некоторых простейших (инфузории) половой процесс — конъюгация происходит путем слияния не гамет, а слиянием генеративных ядер из разных клеток. При процессе копуляции сливающиеся гаметы могут быть одинаковыми по размеру и форме (изогамия) или разными (гетерогамия). В случае резких различий между гаметами, когда одна из гамет крупная, неподвижная, без жгутиков (оогамета), а другая мелких размеров, со жгутиками, такая копуляция получила название оогамии. При этом макрогамета (оогамета) приравнивается к яйцеклетке многоклеточных, а микрогамета — к спермию.
Жизненный цикл простейших представляет собой циклически повторяющийся отрезок развития вида между двумя одноименными фазами (например, от зиготы до зиготы). Жизненный цикл простейших может характеризоваться только бесполым типом размножения (от деления до деления), или только половым размножением (от зиготы до зиготы), или
чередованием полового и бесполого размножения (метагенез). В дальнейшем будут рассмотрены более подробно различные типы жизненных циклов простейших.
Классификация. Согласно современным концепциям, в протозоологии простейшие подразделены на семь типов:
| Тип Саркомастигофоры (Sarcomastigophora) | — 25 тыс. видов |
| Тип Апикомплексы (Apicomplexa) | — 4800 видов |
| Тип Микроспоридии (Microspora) | — 800 видов |
| Тип Миксоспоридии (Myxozoa) | — 875 видов |
| Тип Инфузории (Ciliophora) | — 7500 видов |
| Тип Лабиринтулы (Labyrinthomorpha) | — 35 видов |
| Тип Асцетоспоровые (Ascetospora) | — 30 видов |
В основу подразделения простейших на типы положены принципы строения их ядерного аппарата, органелл движения, ряда микроструктур, типов размножения и жизненных циклов.
Так, саркомастигофоры характеризуются наличием органелл движения: жгутиков и псевдоподий, ядрами одного типа (за редкими исключениями), половым процессом (если он имеется) по типу копуляции. Апикомплексы, как исключительно паразитическая группа простейших, обладают особым комплексом органелл на переднем (апикальном) конце молодых клеток для проникновения в клетку хозяина. У них отсутствуют органеллы движения, а жгутики имеются только у мужских гамет. У большинства наблюдается половой процесс — копуляция, и у многих образуются из зиготы ооциста со спорами, с молодыми паразитами — спорозоитами.
Микроспоридии — внутриклеточные паразиты, образующие одноклеточные споры с амебоидным зародышем — споробластом и с одной полярной нитью, свернутой спирально внутри споры. При выстреливании полярной нити амебоидный зародыш по каналу нити попадает в клетку хозяина. Затем в зародыше происходит автогамия — слияние ядер.
Миксоспоридии — тканевые паразиты животных, имеющие форму плазмодия с множеством ядер. У них наблюдается ядерный дуализм. Они образуют многоклеточные споры с несколькими полярными капсулами, в каждой из которых находится свернутая спирально полярная нить, половой процесс — автогамия.
Инфузории передвигаются при помощи органелл движения — ресничек или их производных; обладают ядерным дуализмом и полиэнергидностью. Половой процесс осуществляется при помощи конъюгации.
Лабиринтулы обитают на водных морских растениях и представляют собой лабиринт цитоплазматических тяжей, по которым передвигаются веретеновидные клетки. Размножаются зооспорами со жгутиками.
Асцетоспоровые — паразиты с многоклеточными спорами с одним или несколькими споробластами, но без стрекательных капсул.
Сравнительная характеристика типов простейших приведена в таблице 1.
Таблица 1. Сравнительная характеристика типов простейших
Источник



