
- IV. Взаимодействие гормона с клеткой-мишенью
- Мембранный способ рецепции какие гормоны
- ЗНАЧЕНИЕ ВЕГЕТАТИВНЫХ НЕРВНЫХ ВЛИЯНИЙ И АКТИВНОСТИ ГИПОТАЛАМО-ГИПОФИЗАРНОЙ СИСТЕМЫ В РЕГУЛЯЦИИ ФУНКЦИИ ПЕРИФЕРИЧЕСКИХ ЭНДОКРИННЫХ ЖЕЛЕЗ, ОБЩЕСОМАТИЧЕСКОГО И МЕТАБОЛИЧЕСКОГО СТАТУСОВ
- Лекция 2 рецепция и механизмы действия гормонов
- Рецепция гормонов
- Локализация рецепторов и виды вторичных посредников
IV. Взаимодействие гормона с клеткой-мишенью




![]()
![]()
Взаимодействие гормона с клеткой-мишенью включает процесс рецепции и трансдукции.
Рецепция. Основана на существовании на поверхности или внутри клетки специальных веществ (в частности, олигопептидов), обладающих высокой избирательной чувствительностью (аффинностью) к сигнальной молекуле-лиганду.
Трансдукция. Образование комплекса гормон – рецептор инициирует активацию внутриклеточных процессов, обеспечивающих в конечном итоге ответ клетки, обусловленный ее спецификой. Это явление получило название трансдукции биологического сигнала в клеточный ответ.
В зависимости от локализации рецепторов различают два основных типа взаимодействия: мембранный (контактный, внеклеточный) и внутриклеточный, включающий, в свою очередь, два варианта: цитозольный и ядерный
(рис. 2).
1. Мембранный тип рецепции характерен для белковых и полипептидных гормонов, для которых мембрана клеток непроницаема. Последовательность взаимодействий при этом: взаимодействие гормона и рецептора → образование активного комплекса «гормон + рецептор» → активация соответствующего фермента, включающего определенную систему вторичных (вторых) посредников, которые обеспечивают формирование ответа клетки на сигнал (гормон). Известны следующие системы вторых посредников (мессенджеров).
А. Системы циклических нуклеотидов. Последовательность событий: активный комплекс «гормон + рецептор» → активация аденилатциклазы (или гуанилатциклазы) → образование из АТФ (или ГТФ) циклического АМФ (или цГМФ) → активация соответствующих протеинкиназ → фосфорилирование и активация белков клетки. Прекращение эффекта обеспечивается инактивацией циклических нуклеотидов (фермент фосфодиэстераза) и их вымыванием из клетки. Циклический АМФ является вторичным мессенджером для АКТГ, ФСГ, ЛГ, ТТГ, паратгормона, кальцитонина, цГМФ – для окситоцина, атриального натрийуретического пептида и некоторых других гормонов.
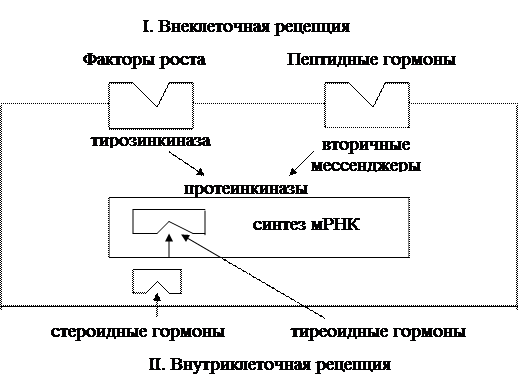
Рис. 2. Виды рецепции клеткой биологических сигналов.
Б. Фосфатидилинозитоловый цикл. Последовательность взаимодействий: активный комплекс «гормон + рецептор» → активация фосфоинозитдиэстеразы (фосфолипазы С) через комплекс с протеином G → гидролиз фосфатидилинозитола 4,5-дифосфата → образование инозитол-1,4,5-трифосфата и диацилглицерола. Дальнейшая организация активного ответа клетки реализуется следующими двумя механизмами:
Ø Инозитол-1,4,5-трифосфат → выход ионов кальция из эндоплазматического ретикулума → образование активного комплекса «кальций + кальмодулин» (рецепторный белок для кальция) → активация «кальций + кальмодулин»-зависимой протеинкиназы → фосфорилирование белков клетки.
Ø Диацилглицерол с участием ионов кальция → активация протеинкиназы С → образование эйкозаноидов и модуляция активности ионных каналов.
В. Тирозиновые протеинкиназы. Схема взаимодействий: образование комплекса «гормон + рецептор» → аллостерическая активация внутриклеточного белкового домена рецептора, обладающего протеинкиназной активностью → аутофосфорилирование тирозинкиназы → фосфорилирование клеточных белков.
2. Внутриклеточный тип рецепции.
А. Цитозольный тип внутриклеточной рецепции. Характерен для стероидных гормонов, проникающих через мембрану клеток. Схема взаимодействия: образование активного комплекса «гормон + рецептор» → транслокация комплекса в ядро клетки и связывание с хроматиновым рецептором → избирательная инициация транскрипции мРНК → изменение белкового синтеза.
Б. Ядерный тип внутриклеточной рецепции установлен для тиреоидных гормонов. Внутриклеточные рецепторы локализованы в ядре клетки и связаны с хроматином. Связывание рецепторов с тиреоидными гормонами усиливает экспрессию генетической информации с усилением синтеза специфической мРНК, кодирующей ферментные белки соответствующего биологического ответа.
Виды ответов клеток на воздействие гормонов
Выделяют следующие проявления воздействия гормонов на клетку-мишень:
1) изменение синтеза и активности ферментов внутри клетки;

2) изменение проницаемости клеточных мембран;
3) влияние на регуляторный аппарат клеток (на число и аффинность рецепторов);
4) модуляция нервных влияний на клетки-мишени.
Основные типы гормональных эффектов
1. Метаболический эффект, состоящий в характерном для каждого гормона изменении обмена веществ. Является практически универсальным.
2. Морфокинетическое действие, проявляющееся в выраженном влиянии гормона на процессы роста и развития тканей (морфогенеза). В наибольшей степени присущ тиреоидным гормонам и гормону роста.
3. Кинетическое (пусковое) действие, состоящее в активации функции, которая без этого влияния не осуществляется или резко ослабевает. Классическим примером этого типа действия служат эффекты тропных гормонов гипофиза.
4. Корригирующее действие проявляется в отношении функции, которая вполне может осуществляться без действия гормона, но им регулируется (пример: регулирующее действие адреналина на функцию сердца).
5. Пермиссивное (разрешающее, позволяющее) действие, состоящее в своей сущности в изменении уровня реактивности клетки и модуляции ее ответа на сигнальное воздействие. Проявляется при подпороговых концентрациях гормона (пример: действие кортизола в отношении адреналина).
Источник
Мембранный способ рецепции какие гормоны

ЗНАЧЕНИЕ ВЕГЕТАТИВНЫХ НЕРВНЫХ ВЛИЯНИЙ И АКТИВНОСТИ ГИПОТАЛАМО-ГИПОФИЗАРНОЙ СИСТЕМЫ В РЕГУЛЯЦИИ ФУНКЦИИ ПЕРИФЕРИЧЕСКИХ ЭНДОКРИННЫХ ЖЕЛЕЗ, ОБЩЕСОМАТИЧЕСКОГО И МЕТАБОЛИЧЕСКОГО СТАТУСОВ

Вегетативная нервная система играет исключительно важную роль в регуляции функциональной активности внутренних органов и тканей, направленную, в частности, на поддержание метаболического гомеостаза. Как известно, высшие вегетативные центры парасимпатической нервной системы расположены в переднем гипоталамусе, а симпатоадреналовой системы – в заднем гипоталамусе. Активность этих центров регулируется нисходящими корковыми влияниями, а также восходящей афферентацией с различных рецепторных зон: болевой, термической, проприоцептивной чувствительности, с осмо-, хемо-, волуморецепторов.
В ответ на изменение центрогенных или периферических вегетативных нервных влияний возникают, в одних случаях, достаточно стереотипные нарушения метаболического гомеостаза, а в других – специфические расстройства.
Так, усиление адренергических влияний на различные органы и системы приводит к подавлению секреторной активности слюнных желез, желудка, кишечника, поджелудочной железы. Одновременно возможны спазм сосудов и развитие ишемии почек, активация ренин-ангиотензин-альдостероновой системы, усиление процессов дистальной реабсорбции натрия, экскреции калия, активация факультативной реабсорбции воды при одновременном ограничении процессов фильтрации в почках. Спазм сосудов кожи при активации симпатоадреналовой системы сочетается, как правило, с ограничением потоотделения.
Таким образом, усиление центрогенных и периферических адренергических влияний закономерно сопровождается комплексом типовых реакций адаптации, направленных на усиление задержки воды и натрия в организме. Наоборот, при адекватном степени раздражителя усилении холинергических влияний, сочетающимся с вазодилатирующим эффектом, как правило, возникает активация секреторной способности желудочно-кишечного тракта, поджелудочной железы, печени, подавление реабсорбции натрия и воды в указанных органах и тканях, что способствует усилению экскреции жидкости и электролитов. Чрезмерная активация холинергических влияний на желудочно-кишечный тракт в условиях локального воспаления приводит к усилению перистальтики, двигательной активности кишечника, сокращению времени пребывания пищевых масс в кишечнике и, соответственно, потере воды и электролитов с каловыми массами, развитию выделительного ацидоза.
Вегетативная нервная система находится в тесном взаимодействии со структурами диэнцефальной зоны, в частности с гипоталамусом и таким образом реализует свое влияние на метаболический статус за счет изменения интенсивности освобождения гормонов различными железами внутренней секреции.
Касаясь особенностей структуры и функции диэнцефальной области мозга, следует отметить, что гипоталамус – подбугорная область, часть межуточного мозга, расположенная около третьего желудочка, подразделяется на передний, средний и задний гипоталамус.
Основными структурами переднего гипоталамуса являются nucleus supraopticus, супрахиазматическая область, nucleus paraventricularis.
Структуры средней части гипоталамуса включают nucleus ventromedialis nucleus dorsomedialis, а задняя часть гипоталамуса включает nucleus hypothalami posterior.
Гипоталамус является местом интеграции не только вегетативных нервных влияний, но содержит терморегулирующий, осморегулирующий центры и центры сытости.
Одной из ведущих функций гипоталамуса является нейроэндокринная, включающая продукцию гормонов вазопрессина и окситоцина, а также либеринов и статинов. Последние играют регулирующую роль (активирующую и ингибирущую) в продукции гормонов аденогипофизом.
Гистологически в аденогипофизе различают три группы клеток: эозинофильные, базофильные (хромофобные), а в соответствии с функциональными особенностями – пять типов клеток: соматотропы, лактотропы, кортикотропы, тиротропы, гонадотропы.
Соматотропы составляют 50% клеток аденогипофиза и продуцируют соматотропный гормон (СТГ). Лактотропы, продуцируют пролактин и составляют 15-20%. Тиреотропный гормон (ТТГ), продуцируемый тиреотропами клеток аденогипофиза, включает около 5% клеток аденогипофиза. На долю кортикотропов, продуцирующих АКТГ, приходится около 15-20% клеток аденогипофиза, а гонадотропы составляют около 10-15% клеток передней доли гипоталамуса.
Гормоны аденогипофиза в зависимости от осбенностей структуры подразделяются на три группы:
1) белки (СТГ и пролактин);
2) гликопротеиды (ФСГ, ЛГ, ЛТГ);
3) полипептиды (АКТГ, липотропины, МСГ, эндорфины).
В то же время в гипоталамусе индентифицировано освобождение соматостатина, пролактостатина, меланостатина.
Процесс высвобождения тропных гормонов аденогипофиза стимулируют релизинг факторы гипоталамуса, включающие люлиберин, кортиколиберин, соматолиберин, пролактолиберин, тиролиберин, меланолиберин..
Регулирующее влияние гипоталамо-гипофизарной системы на периферические эндокринные железы и, соответственно, на процессы обмена веществ находятся под модулирующим влиянием центральной нервной системы, лимбической системы и ретикулярной формации.
В регуляции секреции гормонов гипофиза принимают участие и многие гормоны, продуцируемые клетками АПУД-системы, т. е. эндокринными клетками, рассеянными в различных органах и тканях. К этой группе регуляторов активности гипоталамо-гипофизарной системы относятся субстанция Р, вазоактивный интестинальный пептид, секретин, кальцитонин, ангитензин I и II, предсердный натриуретический фактор и ряд других соединений.
Биологические эффекты указанных гормонов осуществляются в процессе их взаимодействия с соответствующими рецепторами. Для белковых гормонов, факторов роста, нейротрансмиттеров, катехоламинов, простагландинов рецепторы расположены на клеточных мембранах. В связи с этим модификация структуры мембраны под влиянием различных патогенных факторов, формирование «атипизма» рецепторов приводят к «ускользанию» клеток от нервных, гуморальных и гормональных влияний.
Биологические эффекты гормонов с мембранным типом рецепции опосредуются вторичными мессенджерами (ц АМФ, ц ГМФ, инозитолфосфатом, ионизированным кальцием, диацилглицерином и другими).
Гормоны стероидной природы – андрогены, эстрогены, глюко- и минералокортикоиды, кальцитриол, а также тиреоидные гормоны имеют внутриклеточный рецептор в связи с их липофильностью и способностью достаточно легко проникать через липидный бислой цитоплазматических мембран. Гормоны с внутриклеточным типом рецепции проникают в ядро, вызывают дерепрессию определенных участков генома с последующим усилением процессов транскрипции и трансляции соответствующих белков-ферментов. Количество клеточных рецепторов постоянно меняется, отражая функциональную активность клеток и в то же время находится в обратной взаимосвязи с концентрацией гормона в крови и тканях.
В связи с вышеизложенным, этиологические факторы активации или подавления вегетативных, нервных и нейроэндокринных влияний на функциональные системы организма, поддерживающие метаболический и, в частности, водно-солевой гомеостаз, чрезвычайно гетерогенны. Они могут носить врожденный или приобретенный характер, иметь инфекционную или аллергическую природу, возникать как следствие нарушения микрогемодинамики в структурах мозга, воздействия стрессорных раздражителей или онкогенных факторов.
Тесная морфологическая и функциональная взаимосвязь ретикулярной формации ствола мозга, гипоталамических структур и жизненно-важных центров продолговатого мозга может проявляться в условиях патологии стереотипным комплексом так называемых гипоталамических или диэнцефальных синдромов.
Гипоталамические синдромы в соответствии с данными литературы можно представить в виде нескольких групп:
1) нейроэндокринных и нейротрофических расстройств;
2) вегетативно-сосудистых нарушений;
3) мышечно-дистрофических и двигательных расстройств;
4) расстройств психоэмоциональной сферы, в виде нарушений сна и бодроствования.
В данном цикле лекций сделан акцент на ряде нейро-эндокринных нарушений функции гипоталамо-гипофизарно-надпочечниковой системы, периферических эндокринных желез и внутренних органов, приводящих к различным метаболическим расстройствам, в частности сдвигам водно-электролитного гомеостаза.
Источник
Лекция 2 рецепция и механизмы действия гормонов
Рецепция гормонов
Гормоны током крови разносятся по всему организму, попадают во все органы, но действуют только на те из них, которые чувствительны к данному гормону. В этих органах – мишенях имеются специфические воспринимающие структуры – рецепторы, которые трансформируют гормональный сигнал в гормональное действие. Одни рецепторы располагаются поверхностно на мембранах, другие располагаются внутриклеточно.
В 50% случаев гормональные рецепторы локализуются на мембранах клеток-мишеней, в остальных случаях – внутри клетки. Гормоны, которые не способны проникать через плазматическую мембрану, имеют рецепторы на поверхности клетки. Такие рецепторы имеют белковые гормоны, а также катехоламины, простагландины, нейротензин, вещество Р, гистамин.
Стероидные и тиреоидные гормоны воспринимаются внутриклеточными рецепторами.
Ко многим гормонам рецепторы еще не выявлены.
Локализация рецепторов и виды вторичных посредников
Рецепторы обладают высоким сродством и избирательностью к гормонам. Однако они могут связывать и структурные аналоги гормонов. Одни из них могут связываться с рецепторами и вызывать действие, аналогичное действию самого гормона (это агонисты или миметики). Другие – связываются с рецепторами, но при этом не вызывают биологического эффекта или мешают связыванию с гормоном (это антагонисты или литики). Все связывающиеся с данным рецептором соединения называются его лигандами.
Рецепторы являются белковыми структурами. На мембране одной и той же клетки может располагаться большое число различных рецепторов.
Гормональные рецепторы плазматических мембран – это один из видов многочисленных мембранных молекул.
β- Рецепторы катехоламинов принадлежат к семейству, члены которого имеют по семь внутримембранных доменов, а также внутриклеточный белоксвязывающий домены (рис.2).
Рис. 2. Типичный мембранный рецептор.
Трансмембранные, внеклеточные и внутриклеточные домены обладают специфическими связывающими особенностями.
Вначале гормон образует со своим рецептором комплекс, который впоследствии взаимодействует с так называемыми G-белками (белками, связывающими гуаниловые нуклеотиды) клеточной мембраны. Комплекс гормон – рецептор подвергается конформационным изменениям, которые придают ему способность взаимодействовать с этими белками. Одни из них (Gс) стимулируют, а другие (GII) ингибируют превращение гуанрозинтрифосфата в гуанозиндифосфат. Каждый G-белок состоит из трех субъединиц – α, β и γ. При активации эти субъединицы диссоциируют, причем α-субъединица (в зависимости от того, образовалась ли она из Gс — или GII –белка) соответсвенно либо стимулирует, либо ингибирует каталитическую единицу. Каталитичпские единицы представляют собой ферменты, такие как аденилатциклаза, гуанилциклаза, фосфолипаза С мили различные протеинкиназы (рис.3).
Рис. 3. Связывание гормона с его мембранным рецептором.
Внутриклеточный домен рецептора сцеплен с G-белком (состоящим из α, β и γ субъединин) и его каталитической единицей, через которую активируются различные внутриклеточные процессы (ЭЦП и ИЦП – соответсвенно экста- и интрацеллюлярное пространство)
Рецепторы тех гормонов, которые проникают в клетку путем диффузии (как стероиды) или другими способами (например, путем эндоцитоза или с помощью специфических молекул-переносчиков), локализуются внутри клетки.
Стероидные рецепторы, как правило, расположены в ядре. Ядерные рецепторы образуют семейства близких белков, функция которых связана с консервативными аминокислотными последовательностями. Так, рецепторы минералокортикоидов, глюкокортикоидов, андрогенов, эстрогенов, прогестерона, йодтиронинов, метаболитов витамина D и ретиноевых кислот входят в одно суперсемейство регуляторных белков.
Любой ядерный рецептор включает ряд компонентов. Области А/В содержат компоненты, специфически активирубющие транскрипцию. Область С высококонсервативна и соответствует ДНК-связывающему домену, содержащему два «цинковых пальца». Область Dой не ясна, примыкает к области Е (также высококонсервативной и присутствующей во всех рецепторах). Эта область активирует транскрипцию и содержит гормонсвязывающий домен. Область F присутсвует не во всех рецепторах.
Ядерные рецепторы стероидов связывают свои гормоны, а затем сами связываются гормончувствительными элементами ДНК своих генов-мишеней, что приводит к активации транскрипции (рис. 4).
Число специфических рецепторных молекул в клетках-мишенях колеблется от 500 до 300 000 и коррелирует со степенью их чувствительности к гормону. Однако в физиологических условиях всего лишь около 5 % рецепторов занята гормоном.
Количество рецепторов одного и того же типа в данной клетке может изменяться, претерпевает суточные и сезонные колебания и зависит от физиологического состояния организма.
Причинами изменений могут явиться:
а) изменение синтеза рецепторов в клетке;
б) изменение уровня гормона в крови (повышение уровня гормона приводит к уменьшению количества рецепторов и наоборот, уменьшение уровня гормона вызывает увеличение количества рецепторов, что приводит к повышению чувствительности клетки);
в) изменение кислотности среды;
г) появление аутоантител к специфическим рецепторам
Источник



