
Мембранный потенциал, механизмы его происхождения. Методы регистрации.
В 1924 г. английский физиолог Донанн установил, что разность потенциалов внутри клетки и вне ее, т.е. потенциала покоя или МП, близка к калиевому равновесному потенциалу. Это потенциал, образующийся на полупроницаемой мембране разделяющий растворы с разной концентрацией ионов калия, один из которых содержит крупные непроникающие анионы. Его расчеты уточнил Нернст. Экспериментально механизмы возникновения разности потенциалов между внеклеточной жидкостью и цитоплазмой установили в 1939 году Ходжкин и Хаксли. Они исследовали гигантское нервное волокно (аксон) кальмара и обнаружили, что внутри клеток имеется избыток калия, а вне их натрия и кальция. Это обусловлено тем, что в клеточную мембрану встроены ионные каналы, регулирующие проницаемость мембраны для ионов натрия, калия, кальция и хлора.
Суммарная проводимость для того или иного иона определяется числом одновременно открытых соответствующих каналов. В состоянии покоя открыты только калиевые каналы и закрыты натриевые. Поэтому мембрана избирательно проницаема для калия и очень мало для ионов натрия и кальция, за счет неспецифических каналов. Ионы калия поступают в цитоплазму и накапливаются в ней. Когда их количество достигает определенного предела, они по градиенту концентрации начинают выходить через открытые калиевые каналы из клетки. Однако уйти от наружной поверхности клеточной мембраны они не могут. Там их удерживает электрическое поле отрицательно заряженных анионов, находящихся на внутренней поверхности. Поэтому на наружной поверхности мембраны скапливаются положительно заряженные катионы калия, а на внутренней отрицательно заряженные анионы. Возникает трансмембранная разность потенциалов.
Выход ионов калия из клетки происходит до тех пор, пока возникший потенциал с положительным знаком снаружи не уравновесит концентрационный градиент калия, направленный из клетки. Т.е. накопившиеся на наружной стороне мембраны ионы калия не будут отталкивать внутрь такие же ионы. Возникает определенный потенциал мембраны, уровень которого определяется проводимостью мембраны для ионов калия и натрия в состоянии покоя.
Так как мембрана в состоянии покоя незначительно проницаема для ионов натрия, необходим механизм выведения этих ионов из клетки. Этот механизм называется натрий-калиевым насосом. Натрий-калиевый насос — это фермент натрий-калиевая АТФ-аза. Его белковые молекулы встроены в мембрану. Он расщепляет АТФ и использует высвобождающуюся энергию для противоградиентного выведения натрия из клетки и закачивания калия в неё. За один цикл каждая молекула натрий-калиевой АТФ-азы выводит 3 иона натрия и вносит 2 иона калия.
В мембране имеются следующие механизмы трансмембранного транспорта ионов и других веществ:
· 1.Активный транспорт. Он осуществляется с помощью энергии АТФ.
· 2.Пассивный транспорт. Передвижение ионов осуществляется по градиенту концентрации без затрат энергии.
· 3.Сопряженный транспорт. Противоградиентный перенос ионов без затрат энергии..
Мембранный потенциал регистрируется с помощью микроэлектродного метода. Для этого через мембрану, в цитоплазму клетки вводится тонкий, диаметром менее 1 мкМ стеклянный микроэлектрод. Он заполняется солевым раствором. Второй электрод помещается в жидкость, омывающую клетки. От электродов сигнал поступает на усилитель биопотенциалов, а от него на осциллограф и самописец (рис).
Потенциал действия, его фазы и механизмы генерации.
Исследования Ходжкина и Хаксли показали, что при возбуждении аксона кальмара возникает быстрое колебание мембранного потенциала, которое на экране осциллографа имело форму пика. Они назвали это колебание потенциалом действия (ПД). Так как электрический ток для возбудимых мембран является адекватным раздражителем, ПД можно вызвать, поместив на наружную поверхность мембраны катод, а внутреннюю анод. При токе пороговой силы МП снижается до критического уровня деполяризации (КУД), при котором начинается генерация потенциала действия.
На кривой потенциала действия выделяют следующие фазы:
· 1.Локальный ответ (местная деполяризация), предшествующий развитию ПД.
· 2.Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше 0. Поэтому мембрана приобретает противоположный заряд — внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала.
· 3.Фаза реполяризации. Она начинается при достижении определенного уровня МП.
· 4.Фаза следовой деполяризации. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мсек.
· 5.Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу, МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мсек.
Возникновение ПД обусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые и медленные натриевые каналы.
Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ). сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации.
Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой.
Следовая гиперполяризация связана с повышенной, после ПД, калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос.
Изменяя проводимость быстрых натриевых и калиевых каналов можно влиять на генерацию ПД, а следовательно на возбуждение клеток. Это используется в клинике.Чем выше скорость распространения ПД по мембране клетки, ткани, тем выше ее проводимость.
Соотношение фаз возбудимости с фазами потенциала действия.
Уровень возбудимости клетки зависит от фазы ПД.
· В фазу локального ответа возбудимость возрастает. Это фазу возбудимости называют латентным дополнением.
· В фазу реполяризации ПД, когда открываются все натриевые каналы и ионы натрия лавинообразно устремляются в клетку, никакой даже сверхсильный раздражитель не может стимулировать этот процесс. Поэтому фазе деполяризации соответствует фаза полной невозбудимости или абсолютной рефрактерности.
· В фазе реполяризации все большая часть натриевых каналов закрывается. Однако они могут вновь открываться при действии сверхпорогового раздражителя. Т.е. возбудимость начинает вновь повышаться. Этому соответствует фаза относительной невозбудимости или относительной рефрактерности.
· Во время следовой деполяризации МП находится у критического уровня, поэтому даже допороговые стимулы могут вызвать возбуждение клетки. Следовательно в этот момент ее возбудимость повышена. Эта фаза называется фазой экзальтации или супернормальной возбудимости.
· В момент следовой гиперполяризации МП выше исходного уровня, т.е. дальше КУД и ее возбудимость снижена. Она находится в фазе субнормальной возбудимости.
Источник
3.Мембранный потенциал покоя. Современные представления о механизме его происхождения. Метод его регистрации.


Для исследования биоэлектрических явлений в клетках применяют микроэлектроды (стеклянные пипетки, наполненные электролитом, с очень тонким – 0,5 мкм – кончиком). В таком микроэлектроде электролит играет роль проводника тока, а стекло – изолятора. Когда кончик микроэлектрода находится в межклеточной жидкости, между ним и индифферентным электродом (находящимся там же) разность зарядов равна нулю. Если микроэлектрод ввести внутрь клетки, то регистрирующая установка мгновенно покажет некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости.
При выведении кончика микроэлектрода из клетки возвратным движением или прокалывание ее насквозь разность потенциалов между электродами скачкообразно исчезает. Разность зарядов между внутренней и наружной сторонами мембраны клетки называют мембранным потенциалом (МП). В покое эта величина варьирует от -9 до -100 мВ в зависимости от вида ткани и называется мембранным потенциалом покоя (МПП). Следовательно, в состоянии покоя клеточная мембрана поляризована. Уменьшение величины МПП называют деполяризацией, увеличение – гиперполяризацией, восстановление исходного значения – реполяризацией мембраны.
4. Потенциал действия, его фазы. Современное представление о механизме его генерации.
Потенциал действия — волна возбуждения, перемещающаяся по мембране живой клетки в процессе передачи нервного сигнала. По сути своей представляетэлектрический разряд — быстрое кратковременное изменение потенциала на небольшом участке мембраны возбудимой клетки (нейрона, мышечного волокна или железистой клетки), в результате которого наружная поверхность этого участка становится отрицательно заряженной по отношению к соседним участкам мембраны, тогда как его внутренняя поверхность становится положительно заряженной по отношению к соседним участкам мембраны. Потенциал действия является физической основой нервного или мышечного импульса, играющего сигнальную (регуляторную) роль.
Потенциал действия развивается на мембране в результате её возбуждения и сопровождается резким изменением мембранного потенциала.
В потенциале действия выделяют несколько фаз:
• фаза быстрой реполяризации;
• фаза медленной реполяризации (отрицательный следовый потенциал);
• фаза гиперполяризации (положительный следовый потенциал).
Фаза деполяризации. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. При деполяризации клеточной мембраны до критического уровня деполяризации (КУД) происходит лавинообразное открытие потенциал чувствительных Na+- каналов. Положительно заряженные ионы Na+ входят в клетку по градиенту концентрации (натриевый ток), в результате чего мембранный потенциал очень быстро уменьшается до 0, а затем приобретает положительное значение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
Фаза быстрой и медленной реполяризации. В результате деполяризации мембраны происходит открытие потенциалчувствительных К+- каналов. Положительно заряженные ионы К+ выходят из клетки по градиенту концентрации (калиевый ток), что приводит к восстановлению потенциала мембраны. В начале фазы интенсивность калиевого тока высока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока снижается и реполяризация замедляется. Усиливает реполяризацию поступление в клетку Ca2+ Фаза гиперполяризации развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+/K+ помпы. Поступление в клетку Cl– дополнительно гиперполяризует мембрану Изменение величины мембранного потенциала во время развития потенциала действия связано в первую очередь с изменением проницаемости мембраны для ионов натрия и калия.

Современные представления о механизме его генерации
Методом фиксации мембранного потенциала удалось измерить токи, текущие через плазмолемму аксона (аксолемму) кальмара и убедиться в том, что в покое ток катионов (К + ) направлен из цитоплазмы в интерстиций, а при возбуждении доминирует ток катионов (Na + ) в клетку. В состоянии «покоя» плазмолемма почти непроницаема для ионов, находящихся в межклеточном пространстве(Na + С1 — и НСОз — ,).
При возбуждении проницаемость для ионов натрия на время, равное нескольким миллисекундам, резко возрастает, а затем снова падает. В результате катионы (ионы Na + ) и анионы (С1 — , НСОз) разобщаются на плазмолемме: Na + входит в цитоплазму, а анионы нет. Поток положительных зарядов в цитоплазму не только компенсирует потенциал покоя, но и превышает его. Возникает так называемый «овершут» (или инверсия мембранного потенциала). Входящий поток натрия — результат его пассивного движения по открывшимся мембранным каналам по концентрационному и электрическому градиентам. Выходящий поток этого катиона обеспечивается калий-натриевой помпой.
5. Виды электрических ответов (электротонический потенциал, локальный ответ, потенциал действия). Механизм их возникновения.
В пpоцессе pазвития возбуждения плазматической мембpаны (изменения ее ионной пpоницаемости и электpического состояния) в зависимости от силы pаздpажителя возникает тpи вида электpических ответов:
1. Электpотонический потенциал
3. Потенциал действия
Электротонический потенциал — это пассивный сдвиг величины мембранного потенциала (МП) при действии подпорогового стимула электрического тока.
1. Возникает в ответ на действие катода постоянного тока по силе воздействия меньше 0,5 поpоговой величины
2. Сопpовождается пассивной, слабо выpаженной электpотонической деполяpизацией за счет «-» заpяда катода (ионная пpоницаемость мембpаны пpактически не изменяется), котоpая наблюдается только во вpемя действия pаздpажителя
3. Развитие и исчезновение потенциала пpоисходит по экспоненциальной кpивой и опpеделяется паpаметpами
4. pаздpажающего тока, а также сопpотивлением и емкостью мембpаны
5. Такой вид возбуждения имеет местный хаpактеp и не может pапpспpостpаняться
Источник
Регистрация потенциала покоя




![]()
![]()
Мембранный потенциал покоя
Kогда клетка находится [Б30] в состоянии физиологического покоя, ее внутренний потенциал, отрицателен по отношению к наружному, условно принимаемому за нуль[Б31] . Этот потенциал называют потенциалом покоя (мембранным потенциалом покоя, трансмембранным потенциалом покоя).
Довольно часто термин «мембранный потенциал» используют как синоним термина «потенциал покоя»[Б32] . На наш взгляд, это недопустимо, поскольку термин «мембранный потенциал» просто необходим для обозначения любого значения трансмембранной разницы потенциала, наблюдаемое как в состоянии покоя клетки, так и при возбуждении, в любое время и в любом состоянии[Б33] .
Как можно обнаружить мембранный потенциал покоя?
1. методом повреждения
2. методом внутриклеточного отведения (рис. ).
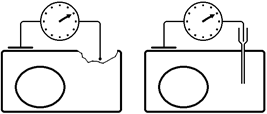
Рис. . Методы регистрации мембранного потенциала покоя: слева – метод повреждения, справа – метод внутриклеточного отведения[Б34] .
Студенты на этот вопрос часто отвечают так: методами внутриклеточного и внеклеточного отведения. Типичная ошибка! Методом внеклеточного отведения можно зарегистрировать возбуждение участка мембраны, сравнив его потенциал с потенциалом мембраны в состоянии покоя, но не потенциал покоя. Но об этом позже.
Метод повреждения чаще используется при регистрации потенциала покоя на макропрепарате – мышце, нерве (рис. ).
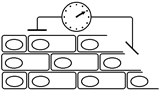
Рис. . Схема регистрации мембранного потенциала покоя методом повреждения на макропрепарате.
Впервые таким способом регистрировали потенциал покоя в середине XIX века Дюбуа-Реймон (на срезе нерва[Б35] ) и Маттеучи (на срезе мышцы). Этот же метод лежит в основе второго опыта Гальвани[Б36] , в этом случае мышечно-нервный препарат выполняет роль регистрирующего прибора.
Методом повреждения достаточно точно измерить потенциал покоя невозможно.
Чтобы измерить потенциал покоя и тем более проследить его изменения, вызываемые каким-либо воздействием на клетку, применяют технику внутриклеточных микроэлектродов (рис. ).
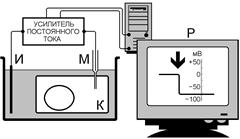
Рис. . Схема измерения потенциала покоя клетки с помощью внутриклеточного электрода.
К – объект исследования – клетка, М – микроэлектрод, И – индифферентный электрод, Р – регистрирующее устройство. Момент прокола на кривой «разность потенциалов – время» показан стрелкой.
Микроэлектрод представляет собой микропипетку, т.е. тонкий капилляр, вытянутый из стеклянной трубочки. Диаметр его кончика около 0,5 мкм[Б37] . Микропипетку заполняют солевым раствором (обычно 3 М KС1), погружают в него металлический электрод (хлорированную серебряную проволочку) и соединяют с электроизмерительным прибором[Б38] , снабженным усилителем постоянного тока[Б39] .
Микроэлектрод устанавливают над исследуемым объектом, например скелетной мышцей, а затем при помощи микроманипулятора — прибора, снабженного микрометрическими винтами, вводят внутрь клетки[Б40] . При удачном введении микроэлектрода мембрана плотно охватывает его кончик и клетка сохраняет способность функционировать в течение нескольких часов, не проявляя признаков повреждения[Б41] .
Микроэлектрод является активным (референтным). Электрод сравнения (индифферентный) обычных размеров погружают в нормальный солевой раствор, в котором находится исследуемая ткань[Б42] .
До прокола мембраны микроэлектродом разность потенциалов между активным и индифферентным электродом равна нулю (рис. и рис. А). Kак только микроэлектрод прокалывает поверхностную мембрану клетки, регистрируется разность потенциалов между поверхностью и содержимым клетки, равная потенциалу покоя клетки[Б43] . На рис. показано, что продвижение микроэлектрода внутри протоплазмы (рис. B, C, D) или изменение положения электрода внутри клетки (рис. E) на показаниях вольтметра не сказываются[Б44] . После прокола (рис. F) микроэлектродом мембраны на противоположной стенке клетки от входа в клетку электрода разность потенциалов вновь не регистрируется. Это свидетельствует о том, что разница потенциал действительно определяется между цитоплазмой и окружающим клетку наружным раствором[Б45] .
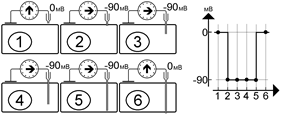
[Б47] Рис. . Результаты измерения разности потенциала микроэлектродным методом при разном расположении активного электрода.
У различных клеток мембранный потенциал покоя варьирует от –50 до –90 мВ[Б48] .
Источник



