
1.2. Механизмы действия гормонов
Гормоны, секретируемые железами внутренней секреции, связываются с транспортными белками плазмы или в некоторых случаях адсорбируются на клетках крови и доставляются к органам и тканям, влияя на их функцию и обмен веществ. Некоторые органы и ткани обладают очень высокой чувствительностью к гормонам, поэтому их называют органами-мишенямиилитканями–мишенями. Гормоны влияют буквально на все стороны обмена веществ, функции и структуры в организме.
Согласно современным представлениям, действие гормонов основано на стимуляции или угнетении каталитической функции определенных ферментов. Этот эффект достигается посредством активации или ингибирования уже имеющихся ферментов в клетках за счет ускорения их синтеза путём активации генов. Гормоны могут увеличивать или уменьшать проницаемость клеточных и субклеточных мембран для ферментов и других биологически активных веществ, благодаря чему облегчается или тормозится действие фермента.
Различают следующие типы механизма действия гормонов: мембранный, мембранно-внутриклеточный и внутриклеточный (цитозольный).
Мембранный механизм. Гормон связывается с клеточной мембраной и в месте связывания изменяет её проницаемость для глюкозы, аминокислот и некоторых ионов. В этом случае гормон выступает как эффектор транспортных средств мембраны. Такое действие оказывает инсулин, изменяя транспорт глюкозы. Но этот тип транспорта гормонов редко встречается в изолированном виде. Инсулин, например, обладает как мембранным, так и мембранно-внутриклеточным механизмом действия.
Мембранно-внутриклеточный механизм. По мембранно-внутриклеточному типу действуют гормоны, которые не проникают в клетку и поэтому влияют на обмен веществ через внутриклеточного химического посредника. К ним относят белково-пептидные гормоны (гормоны гипоталамуса, гипофиза, поджелудочной и паращитовидной желез, тиреокальцитонин щитовидной железы); производные аминокислот (гормоны мозгового слоя надпочечников – адреналин и норадреналин, щитовидной железы – тироксин, трийодтиронин).
Функции внутриклеточных химических посредников гормонов выполняют циклические нуклеотиды – циклический 3׳,5׳ – аденозинмонофосфат (цАМФ) и циклический 3׳,5׳ – гуанозинмонофосфат (цГМФ), ионы кальция.
Гормоны влияют на образование циклических нуклеотидов: цАМФ – через аденилатциклазу, цГМФ – через гуанилатциклазу.
Аденилатциклаза встроена в мембрану клетки и состоит из 3-х взаимосвязанных частей: рецепторной (R), представленной набором мембранных рецепторов, находящихся снаружи мембраны, сопрягающей (N), представленной особымN–белком, расположенным в липидном слое мембраны, и каталитической (C), являющейся ферментным белком, то есть собственно аденилатциклазой, которая превращает АТФ (аденозинтрифосфат) в цАМФ.
Аденилатциклаза работает по слудующей схеме. Как только гормон связывается с рецептором (R) и образуется комплекс гормон- рецептор, происходит образовагние комплексаN– белок – ГТФ (гуанозинтрифосфат), который активирует каталитическую (С) часть аденилатцеклазы. Активация аденилатциклазы приводит к образованию цАМФ внутри клетки на внутренней поверхности мембраны из АТФ.
Даже одна молекула гормона, связавшегося с рецептором, заставляет работать аденилатцеклазу. При этом на одну молекулу связавшегося гормона образуется 10-100 молекул цАМФ внутри клетки. В активном состоянии аденилатциклаза находится до тех пор, пока существует комплекс гормон – рецептор. Аналогичным образом работает и гуанилатциклаза.
В цитоплазме клетки находятся неактивные протеинкиназы. Циклические нуклеотиды- цАМФ ицГМФ- активируют пртеинкиназы. Существуют цАМФ- зависимые и цГМФ – зависимые протеинкиназы, которые активируются своим циклическим нуклеотидом. В зависимости от мембранного рецептора, связывающего определенный гормон, включается или аденилатцеклаза, или гуанилатцеклазаи соответственно происходит образование или цАМФ, или цГМФ.
Через цАМФ действует большинство гормонов, а через цГМФ- только окситоцин, тиреокальцитонин, инсулин и адреналин.
При помощи активированных протеинкиназ осуществляется два вида регуляции активности ферментов: активация уже имеющихся ферментов путем ковалентной модификации, то есть фосфолированием; изменение количества ферментного белка за счет изменения скорости его биосинтеза.
Влияние циклических нуклеотидов на биохимические процессы прекращается под влиянием специального фермента- фосфодиэстеразы, разрушающей цАМФ и цГМФ. Другой фермент – фосфопротеидфосфаза – разрушает результат действия протеинкиназы, то есть отщепляет фосфорную кислоту от ферментных белков, в результате чего они становятся неактивными.
Внутри клетки ионов кальция содержится очень мало, вне клетки их больше. Они пступают из внеклеточной среды по кальциевым каналам в мембране. В клетке кальций взаимодействует с кальцийсвязывающим белком калмодулином (КМ). Этот комплекс изменяет активность ферментов, что ведет к изменению физиологический функций клеток. Через ионы кальция действуют гормоны окситоцин, инсулин, простагландин F2α. Таким образом, чувствительность тканей и органов к гормонам зависит от мембранных рецепторов, а специфическое регуляторное влияние их определяется внутриклеточным посредником.
Внутриклеточный (цитозольный) механизм действия. Он характерен для стероидных гармонов (кортикостероидов, половых гормонов – андрогенов, эстрогенов и гестагенов). Стероидгные гормоны взаимодействуют с рецепторами, находящимися в цитоплазме. Образовавшийся гормнон-рецепторный комплекс переносится в ядро и действует непосредственно на геном, стимулируя или угнетая его активность, т.е. действует на синтез ДНК, изменяя скорость транскрипции и количество инфармационной (матричной) РНК (мРНК). Увеличение или уменьшение количества мРНК влияет на синтез белка в процессе трансляции, что приводит к изменению функциональной активности клетки.
Источник
10.3. Механизмы действия гормонов
Различают несколько механизмов реализации гормонального сигнала для водорастворимых и водонерастворимых гормонов.
Все гормоны оказывают три конечных эффекта:
изменение количества белков и ферментов за счёт изменения скорости их синтеза;
изменение активности имеющихся в клетки ферментов;
изменение проницаемости клеточных мембран.

10.3.1. Цитозольный механизм действия гидрофобных (липофильных) гормоновЛипофильные гормоны способны проникать в клетку через клеточную мембрану, поэтому рецепторы для них располагаются внутриклеточно в цитозоле, на митохондриях, на поверхности ядра. Рецепторы гормонов чаще всего включают 2 домена: для связывания с гормоном и для связывания с ДНК. Рецептор при взаимодействии с гормоном изменяет свою структуру, освобождается от шаперонов, в результате чего гормон — рецепторный комплекс приобретает способность проникать внутрь ядра и взаимодействовать с определёнными участками ДНК. Это, в свою очередь, ведёт к изменению скорости транскрипции (синтез РНК), а вследствие этого меняется и скорость трансляции (синтез белка).
10.3.2. Мембранный механизм действия водорастворимых гормонов
Водорастворимые гормоны не способны проникать через цитоплазматическую мембрану. Рецепторы для данной группы гормонов располагаются на поверхности клеточной мембраны. Поскольку гормоны не проходят внутрь клеток, между ними и внутриклеточными процессами необходим вторичный посредник, который передаёт гормональный сигнал внутрь клетки. В качестве вторичных посредников могут служить инозитолсодержащие фосфолипиды, ионы кальция, циклические нуклеотиды.
10.3.2.1. Циклические нуклеотиды — цАмф, цГмф — вторичные посредники
Гормон взаимодействует с рецептором и образует гормон — рецепторный комплекс, в котором меняется конформация рецептора. Это, в свою очередь, изменяет конформацию мембранного ГТФ — зависимого белка (G-белка) и ведёт к активации мембранного фермента аденилатциклазы, который переводит АТФ в цАМФ.

Внутриклеточный циклический АМФ служит вторичным посредником. Он активирует внутриклеточные ферменты протеинкиназы, которые катализируют фосфорилирование различных внутриклеточных белков (ферментов, мембранных белков), что приводит к реализации конечного эффекта гормона. Эффект гормона «выключается» под действием фермента фосфодиэстеразы, разрушающей цАМФ, и ферментов фосфатаз, дефосфорилирующих белки.
. 
10.3.2.2. Ионы кальция — вторичные посредники
Взаимодействие гормона с рецептором повышает проницаемость кальциевых каналов клеточной мембраны, и внеклеточный кальций поступает в цитозоль. В клетках ионы Са 2+ взаимодействуют с регуляторным белком кальмодулином. Комплекс кальций-кальмодулин активирует кальцийзависимые протеинкиназы, которые активируют фосфолирирование различных белков и приводят к конечным эффектам.

10.3.2.3. Инозитолсодержащие фосфолипиды —вторичные посредники.
Образование гормон-рецепторного комплекса активирует в клеточной мембране фосфолипазу «С», которая расщепляет фосфатидилинозит на вторичные посредники диацилглицерин (ДАГ) и инозитол-трифосфат (ИФ3). ИФ3активирует выход Са 2+ из внутриклеточных депо в цитозоль. Ионы кальция взаимодействуют с кальмодулином, что активирует протеинкиназы и последующее фосфолирирование белков, сопровождающееся конечными эффектами гормона. ДАГ активирует протеинкиназу «С», которая фосфорилирует сериновые или треониновые специфические белки – мишени, в результате чего может меняться проницаемость мембран, в ряде случаев через систему посредников происходит экспрессия генов.
Источник
Семинар 11
48. Гормоны, понятие, химическая природа, биологическая роль. Общебиологические признаки гормонов.
Гормоны – биологически активные вещества, вырабатываемые в организме специализированными клетками, тканями или органам (железами внутренней секреции) и осуществляющие регуляцию деятельности других органов и тканей, метаболических процессов и физиологических функций организма.
Биологическая роль, контролируют:
1. Рост и развитие организма.
2. Развитие и состояние нервной системы.
3. Половое развитие и функции воспроизводства.
4. Все виды метаболизма.
5. Адаптацию и приспособление.
Общебиологические свойства гормонов:
1. Дистантность дейстия.
2. Высокая биологическая активность.
3. Высокая специфичность регулирующего действия.
4. Опосредованность действия через ферментные системы.
5. Высокая скорость метаболизма.
6. Деятельность гормонов контролируется нервной системой.
Гормоны делятся по химическому строению на три группы:
1. Пептидные (инсулин, окситоцин).
2. Стероидные (альдостерон, тестостерон).
3. Непептидные производные аминокислот (адреналин, норадренали).
49. Понятие о прогормонах. Механизм превращения прогормонов в биологически активные гормоны. Роль прогормонов.
Прогормоны — неактивные формы гормонов, которые легко активируются и переходят в активную форму. Многие гормоны — белки и пептиды, эта форма является предстартовая (резервная) форма для ответа на раздражение от нервной системы.
В виде прогормонов образуются инсулин, паратгормон, липотропин и другие белки. Функциональная роль дополнительной последовательности амииокислот у предшественников гормонов, по-видимому, в каждом случае своя. Например, наличие С-пептида в проинсулине необходимо для правильной укладки в пространстве молекулы в процессе ее биосинтеза, для замыкания соответствующих дисульфидных Связей между будущими цепями А и В инсулина. Значительные размеры С-пептида связаны с тем, что он должен увеличивать растворимость синтезированной молекулы инсулина. После того как вновь синтезированная молекула лроиисулина из-за высокой растворимости диффундирует в цистерны аппарата Гольджи, там происходит отщепление С-пептида ферментом трипсинового типа и образуется уже окончательная форма молекулы — биологически активный инсулин.
Роль прогормонов – РЕЗЕРВНАЯ.
50. Клетки-мишени и клеточные рецепторы гормонов, строение рецептора.
Клетка-мишень — клетки, имеющие специфические рецепторы к данному гормону (рецепторы могут находиться в плазматической мембране, в цитозоле или в ядерной мембране).
Рецепторы к гормонам могут находиться на поверхности клеточных мембран — для пептидных гормонов и адреналин. Для стероидных и тиреоидных гормонов внутри клетки, при этом для одной части (глюкокортикостероидов) в цитозоле, для другой (андрогены, эстрогены) в ядре клетки.
В структуре мембранных рецепторов есть три функц. разных участка.
1. Первый домен – домен узнавания. расположен в N-концевой части полипептидной цепи на внешней стороне мембраны. Обеспечивая узнавание и связывание гормона.
2. Второй домен – трансмембранный. Это может быть как G-белок из 7 а-спиральных полипептидных последовательностей. Либо только одна а-спиральная полипептидная цепь.
3. Третий (цитоплазматический). создает химический сигнал в клетке, который сопрягает узнавание и связывание гормона с определённым внутриклеточным ответом.
Рецепторы стероидных и тиреодиных гормонов состоят из трех функц. областей. С-концевой участокдомен узнавания и связывания гормона.
ЦЕнтральная часть – домен связывания ДНК.
N-концевой участок – домен вариабельной области рецептора, отвечает за связывание с другими белками, вместе с которым участвует в регуляции транскрипции.
51. Механизм передачи регуляторного сигнала в клетку гормоном мембранного способа рецепции.
Гормоны, первичный посредники связываются с рецепторами на поверхности клеточной мембраны, образуют комплекс гормон-рецептор, который трансформирует сигнал первичного посредника в изменение концентрации особых молекул внутри клеткивторичных посредников. Вторичными посредниками могут быть следующие молекулы:
52. Циклическая 3,5 АМФ как посредник между гормонами и внутриклеточными механизмами регуляции. Другие посредники.
Под действие рецептора активизируется фермент аденилатцикалаза, которая катализирует преращение АТФ в цАМФ. Образующая цАМФ активирует протеинкиназу А, которая фосфорилирует другие белки и ферменты.

53. Механизм передачи гормонального сигнала эффекторным системам гормоном цитозольного способа рецепции.
Цитозольный механизм (или ядерный ) свойственен липофильным белкам — стероидам. Они проникают через клеточную мембрану в цитозоль и соединяются с внутриклеточными рецепторами. Комплекс гормон-рецептор проникает в ядро клетки, где избирательно влияет на активность генома, это приводит к снижению или активации синтеза определенных ферментов, что приводит к изменению скорости или направления определенных реакций.
54. Центральная регуляция эндокринной системы. Роль либеринов, статинов, тропных гормонов гипофиза.
В гипоталамических ядрах секретируются регуляторные гормоны, которые поступают в аденогипофиз, где меняется продукция гипофизальных гормонов. Пример: кортиколиберин — активирует синтез АКТГ в гипофизе, пролактостатин — ингибирует синтез пролактина. Либерины —
стимуляторы синтеза гормонов гипофиза, статины — ингибиторы. По химической природе это олигопептиды.
В гипофизе (под регуляцией гипоталамуса) образуются тропные гормоны, которые контролируют функцию периферических желез. В аденогипофизе:
Тиреотропный гормон — гликопротеид, работает по мембранно-клеточному механизму в клетках щитовидной железы (вторичный мессенджер — цАМФ) и в клетках жировой ткани (ускоряет процесс липолиза).
АКТГ (аденокортикотропный гормон) — полипептид из 39 АК. Работает по мембранно-клеточному механизму в клетках коры надпочечников, там он стимулирует реакцию гидроксилирования холистерина, в результате которой образуются кортикостероиды. Кроме того активирует липолиз в жировой ткани.
Гонадотропные гормоны — фолликулостимулирующий — гликопротеид, состоящий из двух субъединиц, лютеинизирующий — также гликопротеид, пролактин — простой белок.
Гормон роста — простой белок. Он стимулирует синтез РНК и белков (анаболическое действие), повышает уровень глюкозы в крови, стимулирует образование гликогена, повышает уровень высших жирных кислот, и оказывает еще ряд физиологических эффектов.
Липотропины a и b — два близких по АК набору белка. Активируют липолиз в клетках жировой ткани. b — липотропин предшественник эндорфинов (опиатоподобные эффекты).
55. Эйкозаноиды, понятие, химическое строение, представители, их роль в регуляции метаболизма и физиологических функций.
Эйкозаноиды – биологически активные вещества, синтезируемые большинством клеток из полиеновых жирных клеток, содержащих 20 углеродных атомов.
Включают в себя простагландины, тромбоксаны, лейкотриены.
Простагландины – 20-ти углеродные жирные кислоты, содержат 5-ти углеродное кольцо и гидрокси и/или кетогруппы. Модулируют действия гормонов, оказывают аутокринное влияние на клеток. Вызывают сокращение гладкой мускулатуры, регулируют приток крови к определенному органу. Контролируют транспорт ионов через мембрану.
Лейкротриены – производные 20-ти углеродных полиненасыщенных кислот. Являются медиаторами воспалительных реакций, способствуют сокращению коронарных сосудов, скоращению мышечной ткань бронхов.
56. Нарушение функций эндокринных желез.
Нарушение выработки соматотропного гормона: Гипофункция: нанизм, карликовость. Гиперфункця: гигантизм, акромегалия.
Недостаток тропных гормонов приводит к снижению функции периферических желез.
Заболевания щитовидной железы. Гипотиреоз у новорожденных – кретинизм. Гипотиреоз – микседема, эндемический зоб. Гипертиреоз – диффузный токсический зоб.
Гипофункция – острая надпочечниковая недостаточность. Гиперфункция – стероидный диабет.
Патология мозгового вещества надпочечников – феохромоцитома.
Заболевание поджелудочной железы Сахарный диабет первого и второго типа.
57. Строение, биосинтез и метаболизм иодтиронинов. Влияние на обмен веществ. Гипо- и гипертиреозы, механизм возникновения и последствия.

Белковый обмен: Усиливает транспорт аминокислот в клетки. Активирует синтез дифференцировочных белков в ЦНС, гонадах, костной ткани и обусловливает развитие этих тканей.
Углеводный обмен: Увеличивает гликогенолиз и аэробное окисление глюкозы.
Липидный обмен: Стимулирует липолиз, β-окисление жирных кислот, подавляет стероидогенез. Нуклеиновый обмен: Активирует начальные стадии синтеза пуринов и синтеза пиримидинов, стимулирует дифференцировочный синтез РНК и ДНК.
Гипотиреоз — состояние, обусловленное длительным, стойким недостатком гормонов щитовидной железы, противоположное тиреотоксикозу. Крайняя степень проявления клинической симптоматики гипотиреоза у взрослых — микседема, у детей — кретинизм.
Гипертиреоз — синдром, обусловленный гиперфункцией щитовидной железы. Проявляющийся повышением гормонов: трийодтиронин, тироксин. Гипертиреоз в зависимости от уровня возникновения нарушения различают: первичный — щитовидная железа, вторичный — гипофиз, третичный — гипоталамус.
58. Инсулин, строение, образование из проинсулина. Влияние на обмен углеводов, липидов, аминокислот.
Инсулин – полипептид, состоящий из двух полипептидных цепей. Цепь А содержит 21 аминокислотных остатков, цепь В – 30 аминокислотных остатков. Обе цепи соединены между собой двумя дисульфидными мостиками. Может существовать в форме мономера, димера, гексамера.
Синтез препроинсулина происходит на полирибосомах, связанных с эндоплазматическим ретикулумом. Препроинсулин проникает в люмен ретикулума, где от него отщепляется лидирующая последовательность — N-концевой фрагмент, содержащий 24 аминокислотных остатка. Образовавшийся проинсулин (86 остатков) перемещается в люмене к аппарату Гольджи, где упаковывается в секреторные гранулы. В аппарате Гольджи и секреторных гранулах происходит превращение проинсулина в инсулин. Образуется инсулин с 51 аминокислотным остатком и 31 С-пептид.
Инсулин – главный анаболический гормон.
Влияет на транспорт глюкозы, при участии белков переносчиков ГЛЮТ-4 Влияние инсулина на метаболизм глюкозы. Стимулирует утилизацию глюкозы в клетках разными
путями. 50% на процесс гликолиза, 30-40% в жиры. 10% накапливается в виде гликогена. Общие результат — снижение концентрации глюкозы в крови. Влияние инсулина на метаболизм глюкозы осуществляется путём повышения активности и количества ключевых ферментов гликолиза: глюкокиназы, фосфофруктокиназы, пируваткиназы.
Влияние инсулина на метаболизм жиров.
В печени и жировой ткани инсулин стимулирует синтез жиров, обеспечивая получение для этого необходимых субстратов из глюкозы. В адипоцитах инсулин активирует ацетил КоА – карбоксилазу и ЛП-липазу.
Влияние на аминокислоты.
Инсулин увеличивает поглощение большинства аминокислот мышцей. Этот эффект отражает сочетанное стимулирующее действие на транспортные механизмы и синтез белка и тормозящее действие на катаболизм белка в мышце. Увеличивает поглощение ак тканями, стимулирует синтез белка, тормозя распад белка, тормозит распад белка, снижает окисление аминокислот.
59. Адреналин и норадреналин, биосинтез, пути метаболизма, механизм регуляторного
Катехоламины адреналин, норадреналин и дофамин синтезируются в мозговом веществе надпочечников, в симпатической нервной системе и в мозге.
Адреналин — конечный продукт биосинтеза катехоламинов. В целом синтез катехоламинов — это сложный биохимический процесс. Схематически это выглядит так: Тирозин → ДОФА → Дофамин → Норадреналин → Адреналин.
Катехоламины прямо или косвенно повышают активность эндокринных желез, стимулируют гипоталамус и гипофиз. При любой напряженной работе, особенно физической, содержание в крови катехоламинов увеличивается. Это приспособительная реакция организма к нагрузке любого рода. И чем более выражена реакция, тем лучше организм приспосабливается, тем быстрее достигается состояние тренированности. При интенсивной физической работе повышение температуры тела, учащение сердцебиения и др. вызвано выделением в кровь большого количества катехоламинов.
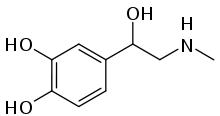
В организме адреналин быстро расщепляется. В отечественных руководствах выделено 3 основных пути обмена адреналина:
1) Окислительное дезаминирование под влиянием моноаминоксидазы с образованием 3, 4- диоксиванилиновой кислоты.
2) Образование ортохинона, превращающегося в адренохром и лейкоадренохром и, в конечном итоге, — в меланин.
3) Образование эфиров серной кислоты.60. Стероидные гормоны (коры надпочечников и половые), структура, метаболизм, механизм регуляторного действия.
61. Принцип качественного открытия гормонов: инсулина, тироксина, 17-кетостероидов.
Для определения инсулина используют качественную реакцию на белковую природу инсулина. Используют 1% раствор сульфата меди и 10% гидроксида натрия. Образуется комплексное соединение с пептидной группировкой.
Метод определения тироксина основан на способности йода, высвободившегося при гидролизе тироксина, давать с крахмалом синее окрашивание.
Обнаружение 17-кетостероидов основано на их способность реагировать в щелочной среде с м-
динитробензолом с образованием продуктов конденсации розового-фиолетового цвета.
62. Программированный контроль–3: знать строение, способ рецепции, механизм действия и биологическую роль тироксина, адреналина, норадреналина, гистамина, серотонина, кортизола, альдостерона, тестостерона, прогестерона, эстрона, эстрадиола, схему строения инсулина.
Воздействует на рецепторы в плазматической мембране. Есть два вида рецепторов: а- адренергические и в-адренергические.
Биологическая роль: регулирует основной обмен, а также усиливает сокращение сердечной мышцы.

Биологическая роль: Норадреналин отличается от адреналина гораздо более сильным сосудосуживающим и прессорным действием, значительно меньшим стимулирующим влиянием на сокращения сердца, слабым действием на гладкую мускулатуру бронхов и кишечника, слабым влиянием на обмен веществ
Воздействует на рецепторы в плазм. мембране: действие норадреналина связано с преимущественным влиянием на α-адренорецепторы.
Биологическая роль: Тироксин влияет на обмен веществ, контролирует рост и развитие организма. Усиливает окислительные процессы в клетках всего организма, в частности и клетках мозга.
в ядре клетки, учавствуя в экспрессии генов.
на плазматической мембране для удержания около клетки.
— стимулирует секрецию желудочного сока, слюны.
— повышает проницаемость капилляров, вызывает отеки, снижает АД.
— вызывает аллергическую реакцию

— является медиатором боли. и др.
Способ рецепции: хрен его знает, скорее всего мембранная.
ГАМК – гамма-аминомаслянная кислота.
Биологическая роль: тормозной медиатор. Способ рецепции: мембранный.
Биологическая роль: стимулирует сокращение гладкой мускулатуры; регулирует АД, температуру, дыхание; обладает антидепрессантным действием.
Способ рецепции мембранный.
Биологическая роль: кортизол является регулятором углеводного обмена организма, а также принимает участие в развитии стрессовых реакций. Для кортизола характерен суточный ритм секреции: максимальная концентрация отмечается в утренние, а минимальная концентрация в вечерние часы.
Способ рецепции: цитозольный и ядерный.
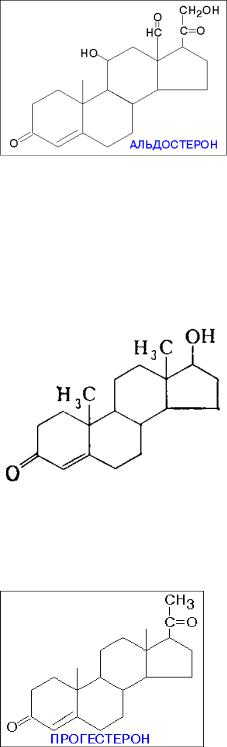
Биологическая роль: минералокортикоиды вызывают усиление канальцевой реабсорбции катионов натрия, анионов хлора и одновременно усиливают канальцевую экскрецию катионов калия и повышают гидрофильность тканей (способность тканей удерживать воду), способствуют переходу жидкости и натрия из сосудистого русла в ткани.
Способ рецепции: цитозольный и ядерный.
Биологическая роль: формирование первичных и вторичных половых признаков. Способ рецепции: ядерный.
Биологическая роль: воздействует на репродуктивную функцию организма, также на ЦНС. Способ рецепции: цитозольный, ядерный.
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Источник



