
- Костная ткань растет способом только оппозиционным
- Костная ткань растет способом только оппозиционным
- Стадии развития скелета в филогенезе.
- Развитие скелета в онтогенезе у человека.
- Первичные и вторичные кости.
- Характеристика остеобластов и остеокластов развиваюшейся кости.
- Cпособы развития костей (окостенения).
- Развитие костей туловища (общие свойства). Развитие и аномалии развития позвонков.
- Развитие и аномалии развития ребер и грудины.
- Развитие костей конечностей.
- Варианты и аномалии развитие костей конечностей.
- Развитие костей черепа.
- Варианты и аномалии развития костей черепа.
- Филогенез соединений костей
- Онтогенез соединений костей
Костная ткань растет способом только оппозиционным
5. Гистогенез и регенерация костной ткани
Развитие костной ткани у эмбриона осуществляется двумя способами:
1) непосредственно из мезенхимы (прямой остеогенез ),
2) из мезенхимы на месте ранее развившейся хрящевой модели кости (непрямой остеогенез ). Постэмбриональное развитие костной ткани происходит при физиологической и репаративной регенерации.
В процессе развития костной ткани образуется костный дифферон : стволовые, полустволовые клетки ( преостеобласты ), остеобласты (разновидность фибробластов), остеоциты. Вторым структурным элементом являются остеокласты (разновидность макрофагов), развивающиеся из стволовых клеток крови.
Прямой остеогистогенез . Такой способ остеогенеза характерен для развития грубоволокнистой костной ткани при образовании плоских костей, например покровных костей черепа. Этот процесс наблюдается в основном в течение первого месяца внутриутробного развития и характеризуется образованием сначала первичной «перепончатой», остеоидной костной ткани с последующей импрегнацией (отложением) солей кальция, фосфора и др. в межклеточном веществе. В первой стадии – образование скелетогенного островка – в местах развития будущей кости происходят очаговое размножение мезенхимных клеток и васкуляризация скелетогенного островка (рис. 42). Во второй стадии, заключающейся в дифференцировке клеток островков, образуется оксифильное межклеточное вещество с коллагеновыми фибриллами – органическая матрица костной ткани (остеоидная стадия). Разрастающиеся волокна раздвигают клетки, которые, не теряя своих отростков, остаются связанными друг с другом. В основном веществе появляются мукопротеиды ( оссеомукоид ), цементирующие волокна в одну прочную массу. Некоторые клетки, дифференцирующиеся в остеоциты, уже в этой стадии могут оказаться включенными в толщу волокнистой массы. Другие, располагающиеся по поверхности, дифференцируются в остеобласты. В течение некоторого времени остеобласты располагаются по одну сторону волокнистой массы, но вскоре коллагеновые волокна появляются и с других сторон, отделяя остеобласты друг от друга. Постепенно эти клетки оказываются «замурованными» в межклеточном веществе, теряют способность размножаться и превращаются в остеоциты. В то же время из окружающей мезенхимы образуются новые генерации остеобластов, которые наращивают кость снаружи (аппозиционный рост).
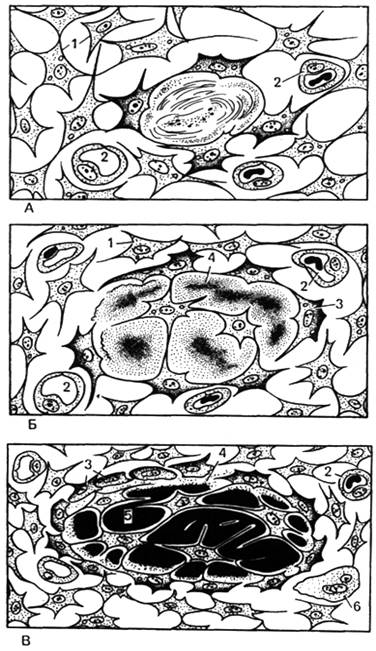
Рис. 42. «Прямой», перепончатый, остеогенез .
А — остеогенный островок (схема); Б — остеоидная стадия (схема); В — оссификация межклеточного вещества (схема); Г — «прямой» остеогенез в плоской кости (микрофотография); 1 — мезенхимные клетки; 2 — кровеносные капилляры; 3 — остеобласты; 4 — остеоид ; 5 — минерализованное межклеточное вещество; 6 — остеокласт.
Третья стадия – кальцификация (импрегнация солями) межклеточного вещества. При этом остеобласты выделяют фермент щелочную фосфатазу, расщепляющую содержащиеся в периферической крови глицерофосфаты на углеводные соединения (сахара) и фосфорную кислоту. Последняя вступает в реакцию с солями кальция, который осаждается в основном веществе и волокнах сначала в виде соединений кальция, формирующих аморфные отложения (Са3(РО 4 )2), в дальнейшем из него образуются кристаллы гидроксиапатита (Са10 (РО4)6 (ОН)2).
Кальцификацию оссеоида связывают с матриксными везикулами.
Процесс биологической минерализации протекает в 2 фазы.
I фаза заключается в образовании исходных кристаллов гидроксиапатита внутри матриксных везикул. Эта фаза контролируется фосфатазами (включая щелочную фосфатазу), а также кальцийсвязывающими молекулами (фосфолипидами и белка ми), которыми матриксные везикулы богаты.
II фаза состоит в разрыве мембран матриксных везикул с выходом сформированных кристаллов в экстрацеллюлярное пространство, где дальнейшее размножение их контролируется условиями внеклеточного микроокружения. Важную роль имеют протеазы и мембранные фосфолипазы , которые обеспечивают разрыв мембран и выход минералов наружу.
Одним из посредников кальцификации является остеонектин – гликопротеин, избирательно связывающий соли кальция и фосфора с коллагеном. В результате кальцификации образуются костные перекладины, или балки. Затем от этих перекладин ответвляются выросты, соединяющиеся между собой и образующие широкую сеть. Пространства между перекладинами оказываются занятыми соединительной волокнистой тканью с проходящими в ней кровеносными сосудами.
К моменту завершения гистогенеза по периферии зачатка кости в эмбриональной соединительной ткани появляется большое количество волокон и остеогенных клеток. Часть этой волокнистой ткани, прилегающей непосредственно к костным перекладинам, превращается в периост, который обеспечивает трофику и регенерацию кости. Такая кость, появляющаяся на стадиях эмбрионального развития и состоящая из перекладин ретикулофиброзной костной ткани, называется первичной губчатой костью. В более поздних стадиях развития она заменяется вторичной губчатой костью взрослых, которая отличается от первой тем, что построена из пластинчатой костной ткани ( четвертая стадия остеогенеза ).
Развитие пластинчатой костной ткани тесно связано с процессом разрушения отдельных участков кости и врастанием кровеносных сосудов в толщу ретикулофиброзной кости. В этом процессе как в период эмбрионального остеогенеза , так и после рождения принимают участие остеокласты.
Костные пластинки обычно образуются вокруг кровеносных сосудов путем дифференцировки прилегающей к ним мезенхимы. Над такими пластинками образуется слой новых остеобластов и возникают новые пластинки. Коллагеновые волокна в каждой пластинке ориентированы под углом к волокнам предыдущей пластинки. Таким образом, вокруг сосуда формируются как бы костные цилиндры, вставленные один в другой (первичные остеомы). С момента появления остеонов ретикулофиброзная костная ткань перестает развиваться и заменяется пластинчатой костной тканью. Со стороны надкостницы формируются общие, или генеральные, пластинки, охватывающие всю кость снаружи. Так развиваются плоские кости. В дальнейшем образовавшаяся в эмбриональном периоде кость подвергается перестройке: разрушаются первичные остеоны и развиваются новые генерации остеонов. Такая перестройка кости практически продолжается всю жизнь.
В отличие от хрящевой ткани кость всегда растет способом наложения новой ткани на уже имеющуюся, т.е. путем аппозиции, а оптимальное кровоснабжение необходимо для дифференцировки клеток скелетогенного островка.
Непрямой остеогистогенез . На 2-м месяце эмбрионального развития в местах будущих трубчатых костей закладывается из мезенхимы хрящевой зачаток, который очень быстро принимает форму будущей кости (хрящевая модель), в виде эмбрионального гиалинового хряща, покрытого надхрящницей. Некоторое время он растет за счет клеток, образующихся со стороны надхрящницы, и за счет размножения клеток во внутренних участках.
Процесс образования кости внутри хрящевого зачатка получил название эндохондрального или энхондрального окостенения (греч. endon — внутри).
Развитие кости на месте хряща, т.е. непрямой остеогенез , начинается в области диафиза ( перихондральное окостенение). Образованию перихондральной костной манжетки предшествует разрастание кровеносных сосудов с дифференцировкой в надхрящнице, прилежащей к средней части диафиза, остеобластов, образующих в виде манжетки сначала ретикулофиброзную костную ткань (первичный центр окостенения), затем заменяющуюся на пластинчатую (рис. 43).
Образование костной манжетки нарушает питание хряща. Вследствие этого в центре диафизарной части хрящевого зачатка возникают дистрофические изменения. Хондроциты вакуолизируются , их ядра пикнотизируются , образуются так называемые пузырчатые хондроциты . Рост хряща в этом месте прекращается. Удлинение перихондральной костной манжетки сопровождается расширением зоны деструкции хряща и появлением остеокластов, которые очищают пути для врастающих в модель трубчатой кости кровеносных сосудов и остеобластов. Это приводит к появлению очагов энхондрального окостенения (вторичные центры окостенения). В связи с продолжающимся ростом соседних неизмененных дистальных отделов диафиза хондроциты на границе эпифиза и диафиза собираются в колонки, направление которых совпадает с длинной осью будущей кости.
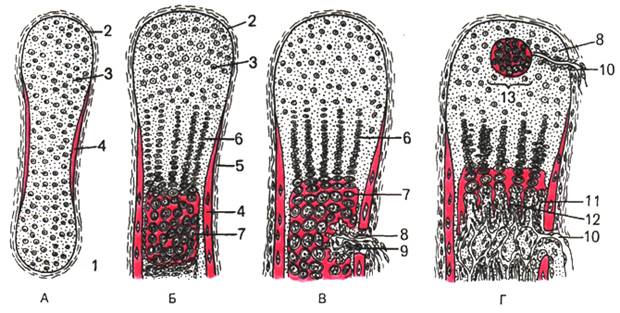
Рис. 43. Непрямой (хрящевой) остеогенез (схема). Образование хрящевой модели кости и перихондральной костной манжетки (по Ю.И.Афанасьеву ).
А, Б, В, Г — стадии остеогенеза ; 1 — первичная хрящевая модель трубчатой кости; 2 -надхрящница; 3 — хрящевая ткань; 4 — перихондральная костная манжетка; 5 — надкостница; 6 — колонки хрящевых клеток; 7 — зона пузырчатых клеток; 8 — врастающая в хрящ мезенхима с дифференцирующимися остеокластами (9) и кровеносными капиллярами (10); 11 — остеобласты; 12 — эндохондрально образованная костная ткань; 13 — точка окостенения в эпифизе.
Таким образом, в колонке хондроцитов имеются два противоположно направленных процесса – размножение и рост в дистальных отделах диафиза и дистрофические процессы в его проксимальном отделе. Одновременно между набухшими клетками происходит отложение минеральных солей, обусловливающее появление резкой базофилии и хрупкости хряща.
С момента разрастания сосудистой сети и появления остеобластов надхрящница перестраивается, превращаясь в надкостницу. В дальнейшем кровеносные сосуды с окружающей их мезенхимой, остеогенными клетками и остеокластами врастают через отверстия костной манжетки и входят в соприкосновение с обызвествленным хрящом. Под влиянием ферментов, выделяемых остеокластами, происходит растворение ( хондролиз ) обызвествленного межклеточного вещества. Диафизарный хрящ разрушается, в нем возникают удлиненные пространства, в которых «поселяются» остеоциты, образующие на поверхности оставшихся участков обызвествленного хряща костную ткань.
Одновременно с процессом развития энхондральной кости появляются и признаки ее разрушения остеокластами. Вследствие разрушения энхондральной костной ткани образуются еще большие полости и пространства (полости резорбции) и, наконец, возникает костномозговая полость. Из проникшей сюда мезенхимы образуется строма костного мозга, в которой поселяются стволовые клетки крови и соединительной ткани. В это же время по периферии диафиза со стороны надкостницы нарастают все новые и новые перекладины костной ткани, образующейся из надкостницы.
Разрастаясь в длину по направлению к эпифизам и увеличиваясь в толщину, они образуют плотный слой кости. Дальнейшая организация периостальной кости протекает иначе, чем организация энхондральной костной ткани. Вокруг сосудов, которые идут по длинной оси зачатка кости из прилегающей к ним мезенхимы, на месте разрушающейся ретикулофиброзной кости начинают образовываться концентрические пластинки, состоящие из параллельно ориентированных тонких коллагеновых волокон и цементирующего межклеточного вещества. Так возникают первичные остеоны. Просвет их широк, границы пластинок нерезко контурированы . Вслед за появлением первой генерации остеонов со стороны периоста начинается развитие общих (генеральных) пластинок, окружающих кость в области диафиза. Вслед за диафизом центры окостенения появляются в эпифизах. Этому предшествуют сначала дифференцировка хондроцитов , их гипертрофия, сменяемая ухудшением питания, дистрофией и кальцинацией. В дальнейшем отмечается процесс окостенения, подобный описанному выше. Оссификация сопровождается врастанием в эпифизы сосудов.
В промежуточной области между диафизом и эпифизами сохраняется хрящевая ткань – метафизарный хрящ, являющийся зоной роста костей в длину.
Физиологическая регенерация костных тканей происходит медленно за счет остеогенных клеток надкостницы, эндоста и остеогенных клеток в канале остеона.
Посттравматическая регенерация костной ткани протекает лучше в тех случаях, когда концы сломанной кости не смещены относительно друг друга. Процессу остеогенеза предшествует формирование соединительнотканной мозоли, в толще которой могут образовываться хрящевые отростки (рис. 44). Оссификация в этом случае идет по типу вторичного (непрямого) остеогенеза . В условиях оптимальной репозиции и фиксации концов сломанной кости регенерация происходит без образования мозоли. Но прежде чем начнут строить кость остеобласты, остеокласты образуют небольшую щель между репонированными концами кости. На этой биологической закономерности основано применение травматологами аппаратов постепенного растягивания сращиваемых костей в течение всего периода регенерации.
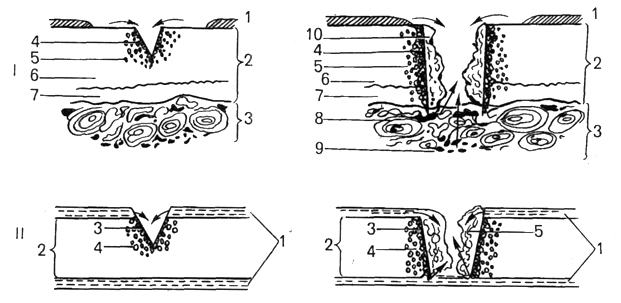
Рис. 44. Посттравматическая регенерация гиалинового хряща при неглубоком (слева) и глубоком (справа) повреждениях.
I — регенерация суставного хряща: I — синовиальная оболочка; 2 — хрящ; 3 — кость; 4 — зона некроза; 5 — зона пролиферации; 6 — некальцифицированный хрящ; 7 — кальцифицированный хрящ; 8 — остеоны с сосудами; 9 — костный мозг; 10 — грануляционная ткань;
II — регенерация реберного хряща: 1 — перихондр; 2 — хрящ; 3 — зона некроза; 4 – зона пролиферации; 5 — грануляционная ткань (по В.Н.Павловой ).
Источник
Костная ткань растет способом только оппозиционным
Лекция «Возрастная анатомия опорно-двигательного аппарата»
Стадии развития скелета в филогенезе.

У животных выделяют наружный и внутренний скелет.
Наружный скелет у разных животных (рис. 1) имеет разное строение и происхождение. У многих беспозвоночных он является продуктом выделения кожного эпителия: кутикула дождевого червя, хитин членистоногих, известковые раковины молюсков.
Наружный скелет у позвоночных появляется в форме чешуи у рыб. Из чешуй у высших рыб развиваются покровные кости головы и плечевого пояса.
Чешуя рыб и кожные окостенения наземных позвоночных всегда дополняются внутренним скелетом.
Внутренний скелет у низших животных (рис. 1) развит слабо и представляет собой систему соединительнотканных образований, иногда включающих рогоподобные волокна, кремниевые или известковые иглы.
Внутренний скелет у головоногих молюсков представлен хрящом.
У позвоночных животных внутренний скелет всегда хорошо развит.
У бесчерепных он перепончатый, у низших рыб – хрящевой, у высших рыб и наземных позвоночных он построен преимущественно из костной ткани.
Развитие скелета в онтогенезе у человека.

Согласно основному биогенетическому закону Геккеля-Мюллера онтогенез есть краткое повторение филогенеза. Онтогенез твердого скелета у человека не является исключением: в развитии костей у человека выделяются три последовательных стадии (рис. 2):
1. Соединительнотканная.
2. Хрящевая.
3. Костная.
Большинство костей в своем развитии последовательно проходят все три стадии – это вторичные кости. Ряд костей при развитии пропускают хрящевую стадию – это первичные кости. К первичным по развитию костям относятся: кости свода черепа, кости лицевого черепа, часть ключицы (акромиальный конец).
Первичные и вторичные кости.

По развитию кости человека делятся на две группы (рис. 3):
- Первичные – проходят в своем развитии две стадии: соединительнотканная и костная.
- Вторичные кости – проходят в своем развитии три последовательных стадии: соединительнотканную, хрящевую и костную.
Характеристика остеобластов и остеокластов развиваюшейся кости.

Для развития костной ткани в костях необходимо наличие популяций двух видов клеток (рис. 4):
- Остеобласты.
- Остеокласты.
Остеобласты представляют собой кубовидной формы клетки (20-30 мкм в диаметре) с одним крупным ядром, располагающиеся близко друг к другу на костном матриксе (межклеточном веществе). Фибробласты продуцируют все компоненты костного матрикса. Они имеют два разных эмбриональных источника:
- нервные гребешковые клетки (выделяются из краев нервного желобка эмбриона при замыкании его в нервную трубку). Они дают начало волокнистой костной ткани костей черепа.
- мезенхимальные клетки закладки кости. Они дают начало пластинчатой костной ткани.
Остеокласты — многоядерные (от 2 до 100 ядер в клетке), большие (от 20 до 100 мкм) клетки гемопоэтической природы. Заносятся в соединительнотканные и хрящевые закладки костей по кровеносным сосудам. Функция остеокластов – резорбция кости.
Для формирования кости как органа необходимо совместная работа двух видов клеток: остеобластов и остеокластов.
Cпособы развития костей (окостенения).


В зависимости от того где начинается формирование костной ткани в костях (включая их закладки) выделяют четыре способа окостенения (рис. 5):
- Эндесмальное окостенение.
- Перихондральной окостенение.
- Энхондральное окостенение.
- Периостальной окостенение.
При эндесмальном окостенении (рис. 5) первичная точка окостенения появляется в центре соединительнотканной закладки кости. Затем новообразующаяся костная ткань распространяется от цента органа к периферии. Таким способом окостеневают первичные кости. На месте первичной точки окостенения обычно наблюдается утолщение (например, теменной бугор, наружный затылочный выступ и т.п.).
Периходральное окостенение характерно для вторичных костей. Остеобласты выстраиваются на поверхности хрящевой закладки кости и начинают синтезировать костный матрикс. Это приводит с сдавливанию и нарушению трофика подлежащей хрящевой ткани, изменения которой активирует остеокласты. В результате этого на поверхности хрящевой закладки кости появляется и постепенно нарастает костная ткань (рис. 5). За счет перихондрального окостенения формируется компактное костное вещество. У длинных трубчатых костей так во внутриутробном периоде образуется диафиз.
При энхондральном окостенении точка (первичный очаг) окостенения появляется в центре хрящевой закладки кости. Затем костная ткань разрастается из центра к периферии (рис. 6). В результате этого формируется губчатое костное вещество. Этим способом развиваются вторичные кости: эпифизы и апофизы трубчатых костей, губчатые, плоские (кроме свода черепа) кости.
Периостальное окостенение происходит за счет надкостницы (periosteum, лат – надкостница). У детей за счет надкостницы кости растут в толщину (напоминаем, что рост кости в длину идет за счет метафизарного хряща)(рис. 6). У взрослых периостальное окостенение обеспечивает физиологическую регенерацию кости.
Развитие костей туловища (общие свойства). Развитие и аномалии развития позвонков.


Рис. 8. Развитие и аномалии развития позвонков.

Рис. 9. Расщелина дуг позвонков на протяжении всех грудных позвонков.
Кости туловища по развитию относятся к вторичным костям. Они окостеневают энхондрально (рис. 7).
Развитие позвонков:
У зародыша закладывается 38 позвонков: 7 шейных, 13 грудных, 5 поясничных, 12-13 крестцовых и копчиковых (рис. 8).
13-й грудной превращается в 1-й поясничный, последний поясничный – в 1-й крестцовый, Идет редукция большинства копчиковых позвонков.
Каждый позвонок имеет первоначально три ядра окостенения: в теле и по одному в каждой половинке дуги. Они срастаются лишь к третьему году жизни.
Вторичные центры появляются по верхнему и нижнему краям тела позвонка у девочек в 6-8 лет, у мальчиков – в 7-9 лет. Они прирастают к телу позвонка в 20-25 лет.
Самостоятельные ядра окостенения образуются в отростках позвонков.
Аномалии развития позвонков (рис. 8, 9):
— Врожденные расщелины позвонков:
— Spina bifida — расщелина только дуг.
— Рахишизис – полная расщелина (тело и дуга).
— Клиновидные позвонки и полупозвонки.
— Платиспондилия – расширение тела позвонка в поперечнике.
— Брахиспондилия – уменьшение тела позвонка по высоте, уплощение и укорочение.
— Аномалии суставных отростков: аномалии положения, аномалии величины, аномалии сочленения, отсутствие суставных отростков.
— Спондилолиз – дефект в межсуставной части дуги позвонка.
— Врожденные синостозы: полный и частичный.
— Os odontoideum – неслияние зуба с телом осевого позвонка.
— Ассимиляция (окципитализация) атланта – слияние атланта с затылочной костью.
— Сакрализация – полное или частичное слияние последнего поясничного позвонка с крестцом.
— Люмбализация – наличие шестого поясничного позвонка (за счет мобилизации первого крестцового).
Развитие и аномалии развития ребер и грудины.

Рис. 10. Развитие и аномалии развития ребер.

Рис. 11. Развитие и аномалии развития грудины.
Развитие ребер (рис. 10):
Закладывается 13 пар ребер. Затем 13-е ребро редуцируется и срастается с поперечным отростком 1-го поясничного позвонка.
Основных точек окостенения в ребре две: точка окостенения на месте будущего угла ребра (окостеневает тело ребра) и в головке ребра (на 15-20 году жизни). У 10 верхних ребер появляется точка окостенения в бугорке ребра.
Передние концы 9 пар верхних ребер образуют грудные полоски – источник развития грудины.
Развитие грудины (рис. 11):
Источником развития грудины являются грудные полоски – расширенные концы хрящевых концов девяти пар верхних ребер. В грудине бывает до 13 точек окостенения.
Аномалии развития ребер (рис. 10):
— Отсутствие ребра
— Отсутствие части ребра
— Дефект ребра
— Раздвоение ребра (вилка Лушки)
— Шейное ребро
— XIII ребро
Аномалии развития грудины (рис. 11):
— Аплазия рукоятки грудины
— Отсутствие отдельных сегментов тела грудины <
— Расщепление грудину
— Отсутствие тела грудины
— Воронкообразная деформация
— Куриная грудь
Развитие костей конечностей.



Рис. 14. Развитие эпифизов трубчатых костей.

Рис. 15. Развитие костей верхней конечности.

Рис. 16. Развитие тазовой и бедренной костей.


Кости конечностей по развитию относятся к вторичным костям. Исключение представляет собой ключица: ее тело и акромиальный конец окостеневают эндесмально (точка окостенения появляется на 6-7-й неделях внутриутробного развития.
Диафизы длинных трубчатых костей окостеневают перихондральными и энходральными способами. В диафизах первичная точка окостенения появляется на 2-м – начале 3-го месяцев внутриутробного развития и растет по направлению к проксимального и дистальному эпифизам.
Эпифизы и апофизы длинных трубчатых костей окостеневают энходральным способом. Они у новорожденных хрящевые. Вторичные точки окостенения появляются в течение первых 5-10 лет жизни. Исключение составляют эпифизы костей, образующих коленный сустав: точка окостенения в дистальном конце бедренной кости появляется на 6 месяце, а в проксимальном конце большеберцовой кости – на 7 месяце внутриутробного развития. Прирастают эпифизы к диафизам после 15-17 лет и позже.
Варианты и аномалии развитие костей конечностей.

Рис. 19. Аномалии развития костей верхней конечности.

Рис. 20. Аномалии развития костей нижней конечности.
Аномалии развития лопатки:
- Глубокая вырезка, иногда отверстие
- Несращение акромиона (синхондроз)
Аномалии развития ключицы:
- Варьируют изгибы
- Нет конусовидного бугорка и трапециевидной линии.
Варианты и аномалии развития плечевой кости
- Processus supracondylaris – над медиальным надмыщелком.
Аномалии развития костей предплечья:
- Локтевой отросток не срастается с телом локтевой кости
- Отсутствие лучевой кости
Аномалии развития костей кисти:
- Добавочные кости запястья, например, os centrale
- Добавочный палец (полидактилия)
Варианты и аномалии развития тазовой кости:
- Отверстие в центре подвздошной ямки
- Удлинение подвздошных остей
Варианты и аномалии развития бедренной кости:
- Увеличение ягодичной бугристости – третий вертел, trochanter tertius
Варианты и аномалии развития костей голени:
- Уплощенное (не трехгранное) тело большеберцовой кости
Варианты и аномалии развития костей стопы
- Добавочные кости предплюсны
- Добавочные пальцы
Развитие костей черепа.


Рис. 22. Источники развития костей лицевого черепа.


Рис. 24. Развитие костей черепа после рождения.
Кости свода и лицевого черепа по развитию относятся к первичным костям, окостеневающим на основе соединительной ткани эндесмальным способом окостенения.
Кости лицевого черепа развиваются на основе жаберных дуг (первой и второй висцеральной дуги).
Из первой висцеральной дуги развиваются следующие кости: верхняя, нижняя челюсти, частично скуловая и небные кости, медиальная пластинка крыловидного отростка клиновидной кости; молоточек, наковальня – слуховые косточки; костное небо и его швы, нижняя часть глазницы.
Из второй висцеральной дуги развиваются: стремечко, шиловидный отросток височной кости, малые рога подъязычной.
Кости основания черепа проходят три стадии развития: соединительнотканную, хрящевую и костную. Т.е. они являются вторичными. Они окостеневают энхондрально.
Варианты и аномалии развития костей черепа.

Рис. 25. Вставочные кости швов черепа (слева), деформации черепа (справа).


Известны следующие аномалии развития черепа
- Непостоянные (вставочные, вормиевы) кости швов
- Кости родничков
- Непостоянные швы
- Большие теменные отверстия
- Тонкая теменная кость
- Дырчатый череп
- Краниосхизис – головной мозг и череп открыты с дорсальной стороны
- Краниостеноз – преждевременный синостоз отдельных или всех швов, ведет к деформациям черепа:
- Башенный
- Ладьевидный
- Клиновидный
- Скошенный
- Микроцефалия
- Макроцефалия
- Гидроцефалия
- Аэнцефалия (отсутствует свод черепа, сочетается с недоразвитием головного мозга).
- Аномалии лицевого черепа:
- Заячья губа
- Волчья пасть
- Колобома
- Др.
Филогенез соединений костей



Рассмотрим филогенез соединения костей хордовых. У беспозвоночных твердого скелета нет, весь скелет представлен соединтельнотканным тяжем — хордой.
У рыб, обитающих в водной среде, многочисленные кости скелета (рис. 28) соединяются при помощи непрерывных соединений: соеденительнотканных и хрящевых.
Важным биомеханическим фактором, повлиявшим на филогенез соединений костей, является выход животных на сушу. Кратковременное пребываниена твердой поверхности (в том числе перемещение с места на место), которое наблюдается, напрмер, у двоякодышащих рыб, приводит к появлению гемиартрозов между костями конечностей (рис. 29). Большинство исследователей считают такую форму пререходной от снартроза к диартрозу.
Окончательный выход животных на сушу формирует два направления морфогенеза мягкого остова. Во-первых, формируются суставы со всеми обязательными и вспомогательными элементами и высокой степенью подвижности. Во-вторых, в местах контакта костей с увеличившейся нагрузкой (из-за возросшего действия силы тяжести) формируются синостозы (кости срастаются между собой). Признаки обоих изменений строения соединений костей уже выявляются у земноводных (рис. 30).
Онтогенез соединений костей

Рис. 31. Варианты дисплазии тазобедренного сустава.
В онтегенезе соединения костей наблюдаются сходные с филогенезом тенденции. Первоначально все соединения образованые скоплением мезенхимальных клеток (эмбриональная соединительная ткань).
В конце первой половины пренатального онтогенеза (16-18-ая недели внутриутробного развития) между костями, которые смещаются (движутся) друг относительно друга, формируются суставы. Важным биомеханическим фактором их морфогенеза являются силы мышц, двигающих кости. Между зачатками костей, которые соединяются без смещание, формируются непрерывные соединения.
У новорожденных имеется закладка всех элементов суставов на нижней конечности. Однако большинство из них достигают функциональной зрелости к юношескому возрасту.
Основной аномалией развития соединения костей является дисплазия суставов. Для данные аномалии характерно изменение формы одной из суставных поверхностей, сопровождающееся изменениями строения расположенных рядом элементов сустава (рис. 31).
Источник



