
Конъюгация как способ генетического картирования бактерий




Глава 3. Картирование генов бактерий
Бактерии — прокариотические микроорганизмы, генетический аппарат которых в основном представлен единственной кольцевой молекулой ДНК, образующей сложную структуру из множества суперспирализованных петель, удерживаемых в компактном состоянии с помощью молекул РНК и белков. Такая скрученная хромосома (folded chromosome), называемая еще ядерным тельцем, или нуклеоидом, связана с мембраной клетки, что является необходимым условием для начала репликации и сегрегации дочерних хромосом в процессе деления клетки. Кроме хромосомы, генетический материал у бактерий может быть представлен также эписомными элементами — профагами и плазмидами, наличие которых не обязательно и нежизненно важно для клетки.
Размер хромосомы бактерий по данным разных методов (генетического: размер трансдуцирующего фрагмента; физических: вискоэластометрический метод, скорость ренатурации, электронная микроскопия) составляет для Pseudomonas aeruginosa 2,1 ⋅ 10 9 Д, для Escherichia coli К-12 — 2,8 ⋅ 10 9 , для Bacillus subtilis — 2,0 ⋅ 10 9 — 2,6 ⋅ 10 9 и для Streptomyces coelicolor — 4,7 ⋅ 10 9 Д. Если исходить из среднего размера гена в 1500 пар нуклеотидов, то бактериальная хромосома может содержать около 3000 генов.
Число хромосом в одной клетке бактерий зависит от стадии развития и физиологических условий роста культуры. При выращивании культуры на богатой среде в условиях хорошей аэрации может быть несколько хромосом в одной клетке вследствие реинициации новых циклов репликации ДНК еще до деления клетки.
Число автономно реплицирующихся кольцевых молекул плазмид определяется системой контроля репликации. При наличии строгого контроля репликации число копий плазмид на одну хромосому невелико, а при ослабленном контроле репликации оно увеличивается на 1-2 порядка, достигая нескольких десятков и сотен копий.
Бактерии как объекты генетических исследований имеют ряд преимуществ перед высшими организмами: они гаплоидны, способны к быстрому и практически неограниченному размножению в довольно простых лабораторных условиях и обладают рядом относительно несложных признаков, удобных для количественного, биохимического и генетического анализов.
В отличие от эукариотических микроорганизмов бактерии обладают разнообразными способами передачи ДНК, ведущими к гетерозиготному состоянию клеток. Почти каждый новый вид микроорганизмов — это новая генетическая система, новый феномен со специфическими особенностями. Не все детали генетического анализа, тщательно разработанные на одной системе, например на Е. coli К-12, в одинаковой степени пригодны для изучения другой системы, например P. aeruginosa или S. coelicolor A3 (3). Следует, однако, отметить, что принципиальные положения или главные приемы, разработанные для классических объектов генетики бактерий, успешно используются для генетического изучения новых групп прокариотических микроорганизмов.
В мире бактерий существует три различных способа переноса генетического материала из одной клетки в другую — конъюгация, трансдукция и трансформация. В последнее время на высших организмах и бактериях успешно и эффективно разрабатывается искусственный способ объединения генетического материала разных клеток путем слияния и регенерации протопластов.
Важное значение для генетического анализа имеют две особенности генетической рекомбинации, обнаруженные у всех исследованных до сих пор бактерий. Первая особенность состоит в относительной редкости генетической рекомбинации в скрещиваемых бактериальных культурах, что, за редким исключением, требует применения селекции рекомбинантов из большой популяции клеток родительских типов. В качестве селектируемых маркеров используют удобные аллели родителей, обусловливающие независимость роста от определенного питательного вещества (прототрофность) или устойчивость к антибиотикам, ядам или вирулентным бактериофагам. Выделенные рекомбинанты затем могут быть классифицированы по отношению к неселективным маркерам путем высева на ряд диагностических сред с добавками факторов роста или отсутствием ингибиторов.
Вторая особенность генетической рекомбинации бактерий, наблюдаемая при каждом из известных в настоящее время способов передачи генетической информации — конъюгации, трансдукции и трансформации, заключается в образовании неполноценных зигот — меродиплоидов — вследствие неодинакового вклада обоих родителей. В то время как генетический материал материнской (реципиентной) клетки представлен в зиготе целой хромосомой, т. е. полным набором генов, материал отцовской (донорной) клетки — только фрагментом хромосомы или частью генома. Образование меродиплоидной зиготы у бактерий сказывается на результатах рекомбинации, обусловливая отсутствие рекомбинантов от единичного или нечетного числа кроссинговеров, так как жизнеспособные рекомбинанты возможны лишь при двойном или четном числе перекрестов между хромосомой реципиента и фрагментом ДНК донорной клетки. Это приводит к уменьшенному вдвое выходу рекомбинантов.
Вклад донора зависит от способа передачи генетического материала, выбора конкретной системы и метода анализа. При трансдукции размер фрагмента хромосомы донора четко ограничен объемом головки бактериофага, в которую может быть ошибочно упакована ДНК хозяина. Например, головка фага Р1 может нести в 2,5 раза больше ДНК, чем головка фага Р22, и в опытах по трансдукции может включать сегмент ДНК Е. coli, равный приблизительно 2,2 % генома клетки, в то время как при генерализованной трансдукции у Salmonella typhimurium фаг может переносить около 1% генома.
При трансформации длина переносимой молекулы ДНК донора очень коротка и зависит как от способа приготовления препарата ДНК, так и от специфики конкретной системы. Размер однонитевой ДНК, участвующей в синапсе при трансформации у В. subtilis, составляет в среднем около 2 МД (около 0,1 % хромосомы). Только при конъюгации размер гетерозиготной области может сильно колебаться и в редких случаях достигать размера полного генома.
Меродиплоидность накладывает определенные ограничения на обнаружение сцепления между генами с помощью измерения частот рекомбинации. Однако данное препятствие можно преодолеть путем применения количественных и качественных тестов сцепления, зависящих только от меродиплоидного состояния и не зависящих от частот рекомбинации между парой локусов. Примером может служить картирование по градиенту переноса маркеров или по времени их вхождения в реципиентную клетку.
Несмотря на перечисленные трудности генетической рекомбинации у бактерий, высокая разрешающая способность методов генетического анализа позволила к настоящему времени картировать с той или иной полнотой хромосомы многих видов бактерий и изучить тонкое строение и функцию отдельных генов.
Источник
Картирование бактериальных генов при конъюгации
При конъюгации происходит постепенный перенос хромосомы до-норной (мужской Hfr) клетки в реципиентную (женскую F-), причем этот процесс может прерваться в любой момент и в реципиентной клетке оказывается лишь та часть хромосомы донора, которая успела к этому моменту в нее проникнуть. Поэтому вероятность рекомбинации между маркерами зависит не только от расстояния между ними, но и от расположения их на хромосоме по отношению к точке начала переноса.
В связи с этой особенностью генетические карты участков бактериальных хромосом могут быть построены при конъюгации двумя независимыми способами: первый способ основан на учете частоты вхождения маркеров, а второй – на учете частоты включения их в реципиентную хромосому, т.е. на учете истинной частоты рекомбинации (кроссинговера).
Первый способ выполняется двумя методами: 1. Картирование по времени переноса маркеров; 2. Картирование по градиенту переноса маркеров. Эти методы пригодны только для ориентировочного картирования (градиент передачи) или для локализации генов, расположенных не менее чем на расстоянии 1-2 мин генетической карты (картирование по времени переноса).
Для тонкого генетического картирования используют второй способ (рекомбинационный), заключающийся главным образом в анализе соотношения частот различных классов рекомбинантов в двухфакторных и трехфакторных скрещиваниях. Этот метод позволяет локализовать гены, которые расположены не далее 3 минут карты, или определить относительное расположение мутаций в пределах одного гена.
Конъюгационные скрещивания проводят путем совместного культивирования штамма-донора со штаммом-реципиентом на плотных или жидких средах. Общим условием конъюгационного скрещивания является выбор признаков донора и реципиента, по которым будет проводиться отбор редко возникающих рекомбинантов из смеси родительских клеток (селективных маркеров), и признаков, по которым будет вестись учет расщепления мерозигот (неселективных маркеров). Причем селективный маркер реципиента принято называть селектором, а селективный маркер донора – антиселектором или контрселектором. По сути, селекция и контрселекция – это просто разные названия одной и той же процедуры. В качестве селективных чаще используют маркеры ауксотрофности и высевают конъюгационную смесь на среду, не содержащую необходимого фактора роста, для контрселекции часто используют антибиотики или фаги, к которым чувствителен донор. Желательно использовать маркеры с низкой частотой реверсий.
Например: в скрещивании Hfr Thr+Leu+Рrо+Str S х F-Thr-Leu — Рго — Str R после высева смеси на минимальную среду с добавлением лейцина (leu), пролина (рrо) и сгрептомицина (str) на ней не вырастут клетки реципиента, так как они нуждаются в отсутствующем в среде треонине (thr), и клетки донора, так как чувствительны к стрептомицину. Вырастут только реципиентные, клетки, получившие маркер Thr+ от донора (рекомбинанты Thr+Str r ). В данном примере признак треонинзависимости реципиента используется в качестве селектора, а в качестве контрселектора – чувствительность донора к стрептомицину. Признаки Leu+ и Pro+, по которым отличаются донор и реципиент, будут неселективными, так как использованная среда не позволяет различать аллели этих генов. Их можно применить для анализа рекомбинационного процесса.
Дата добавления: 2015-02-07 ; просмотров: 2517 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Источник
Конъюгация как способ генетического картирования бактерий
11. Механизмы передачи генетического материала у бактерий: конъюгация, трансдукция, трансформация.
Рекомбинация между геномами бактерий осуществляется 3-мя механизмами: конъюгацией, трансдукцией и трансформацией
Трансформация – передача генетической информации через выделенную из клетки-донора ДНК.
1928г. Ф Гриффит (опыт с вирулентнотью пневмококков)
По происхождению ДНК может быть плазмидной или хромосомной и нести гены, трансформирующие реципиента. Подобным путем среди бактериальных популяций могут распространять гены, кодирующие факторы вирулентности, однако в обмене генетической информацией трансформация играет незначительную роль.
Трансформирующей активностью обладает только двунитчатая высокоспирализованная ДНК.
В клетку-реципиент проникает только одна нить ДНК, другая – в клеточной мембране подвергается деградации с освобождением энергии, необходимой для проникновения в клетку второй нити ДНК.
Интеграция с хромосомой требует наличия гомологичных участков с трансформирующей ДНК.
Процесс трансформации зависит от компетентности клетки-реципиента и состояния трансформирующей донорской ДНК.
Компетентность клеток бактерий (способность воспринимать трансформирующую ДНК) зависит от присутствия в ЦПМ особых белков, обладающих специфическим аффинитетом к ДНК. Компетентность зависит от фазы роста бактериальной культуры, вида бактерий. Может увеличиваться при обработке культуры раствором хлорида кальция при пониженной температуре.
Трансформация служит хорошим инструментом для картирования хромосом, поскольку трансформированные клетки включают различные фрагменты ДНК. Определение частоты одновременного приобретения двух заданных характеристик (чем ближе расположены гены, тем более вероятно, что они оба включатся в один и тот же участок ДНК) дает информацию о взаиморасположении соответствующих генов в хромосоме.
Трансформация является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
Трансдукция – передача бактериальной ДНК посредством бактериофага.
1952г. Дж. Ледебург и Н. Циндер (опыты с сальмонеллами)
Так как трансдуцируются лишь небольшие фрагменты ДНК, вероятность рекомбинации, затрагивающей какой-то определенный признак, очень мала: она составляет от 10 -6 до 10 -8 .
· Общая (неспецифическая) трансдукция – перенос бактериофагом фрагмента любой части бактериальной хромосомы. Феномен неспецифической трансдукции может быть использован для картирования бактериальной хромосомы.
● Специфическая трансдукция – фаговая ДНК интегрирует в бактерию с образованием профага. При исключении ДНК фага из бактериальной хромосомы захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы. Специфическая трансдукция может служить механизмом переноса вирулентных генов среди бактерий при условии, что эти гены локализованы в непосредственной близости от мест интеграции профага.
● Абортивная трансдукция . При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
Обязательное условие – трансмиссивная плазмида ( F , R ), обладающая tra -опероном .
Интегративной трансмиссивной плазмидой является F -фактор . Донорские клетки, имеющие встроенный в хромосому F -фактор, называются Hfr -клетками.
Биологическое значение конъюгации хорошо видно на примере распространения резистентности бактерий к антибиотикам. Устойчивость к антибиотикам бактерия может получить в результате мутации, что происходит 1 раз на каждые 106 клеточных делений. Изменившаяся генетическая информация может быстро распространяться среди сходных бактерий посредством конъюгации.
Источник
Конъюгация как способ генетического картирования бактерий
Длительное время считали, что бактерии — изолированные генетические системы, и каждая особь имеет одного (и только одного) родителя, то есть их изменчивость вызвана лишь мутациями. Никто не мог себе представить, что, подобно гаметам высших организмов, бактерии способны обмениваться генетическим материалом и, по аналогии с половым размножением, давать начало потомству с новыми свойствами. Однако Лёдерберг и Тёйтем (1946), высевая на минеральную среду, предварительно смешав два типа мутантов Escherichia coli (один нуждался в биотине и метионине, другой — в треонине и лейцине), показали, что у бактерий генетические рекомбинанты со способностью к синтезу всех четырёх факторов роста возникают с частотой 10″6, хотя теоретическая вероятность реверсии по двум генам составляет 10-14-10-16 на генерацию. Это противоречие было снято установлением факта прямой передачи генетического материала от донорной клетки к реципиентной (конъюгация). Помимо конъюгации, передача генетического материала у бактерий может осуществляться также с помощью трансформации и трансдукции (рис. 4-16).
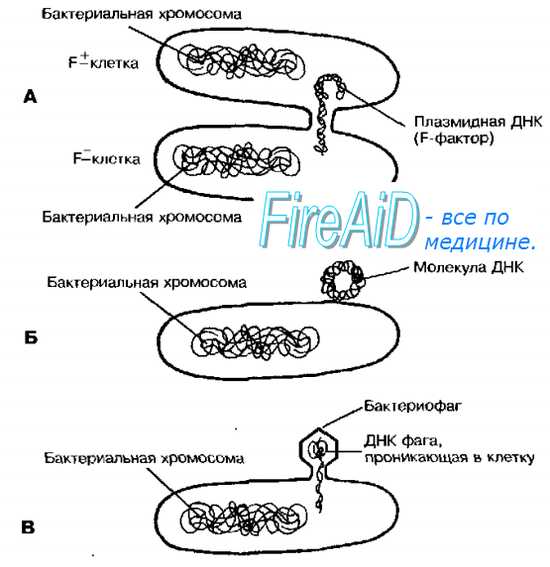 Рис. 4-16. Механизмы перенос бактериальной ДНК. Конъюгация (А), трансформация с использованием отдельной молекулы ДНК (Б) трансдукция с помощью фагов (В).
Рис. 4-16. Механизмы перенос бактериальной ДНК. Конъюгация (А), трансформация с использованием отдельной молекулы ДНК (Б) трансдукция с помощью фагов (В).
Конъюгация бактерий. F-фактор бактерии.
Конъюгация — прямой перенос фрагмента ДНК от донорских бактериальных клеток к реципиентным при непосредственном контакте этих клеток. Биологическая значимость этого процесса стала проясняться после внедрения в медицинскую практику антибиотиков. Устойчивость к антибиотикам можно получить в результате мутации, что происходит один раз на каждые 106 клеточных делений. Однако, однажды изменившись, генетическая информация может быстро распространяться среди сходных бактерий благодаря конъюгации, поскольку каждая третья из близкородственных бактерий способна именно к этому типу генетического переноса. Для реализации процесса необходим F-фактор — плазмида, кодирующая информацию, необходимую для конъюгации.
Конъюгация требует наличия двух типов клеток: доноров (F + ), обладающих F-фактором, и реципиентов (F — ), не обладающих им. При скрещивании клеток F — и F + фактор фертильности передаётся с частотой, близкой к 100%.
Фактор переноса содержит гены специальных и необходимых при конъюгации структур — F-пилей и ряд других генов, вовлечённых в процесс взаимодействия с F — -клетками.
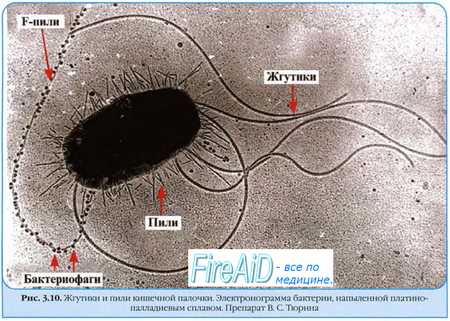
Первый этап конъюгации — прикрепление клетки-донора к реципиенту с помощью F-пилей. Затем между клетками формируется конъюгационный мостик, через который передаётся F-фактор, а также и другие плазмиды, автономно пребывающие в цитоплазме донора. При попадании F-фактора в реципиентную клетку она становится F+ и приобретает способность передавать фактор фертильности другим F — -клеткам. Подобный механизм обеспечивает приобретение популяционно. устойчивости к антибактериальным агентам.
В популяции клеток, содержащих F-плазмиду, только те, в которых она интегрирована в бактериальную хромосому (Hft + -клетки), способны быть донорами хромосомной ДНК. При перенос генетического материала бактериальная ДНК реплицируется, начиная от места включения F-фактора, одна цепь ДНК переносится в реципиентную F’-клетку двигаясь 5′-концом вперёд тогда как другая остаётся в Hfr + -клетке, то есть донор сохраняет своё генетическое постоянство. После начала конъюгации хромосомный материал переносится, начиная от генов, близких к начальной точке транспорта (рис. 4-17).
В бактерии-реципиенты обычно попадают первые из переносимых генов, размер которых зависит от времени, в течение которого проходила конъюгация, и очень редко — все гены. Позже всех переносится участок плазмиды, содержащий ген переноса кодирующий F-пили. Поскольку полная трансмиссия — явление редкое, реципиентная клетка при Hfr-конъюгации обычно остаётся F-. Вслед за процессом переноса в клетке-реципиенте происходит гомологичная рекомбинация между донорской ДНК и собственной ДНК реципиента.
Процесс конъюгации может происходить только при соблюдении ряда условий.
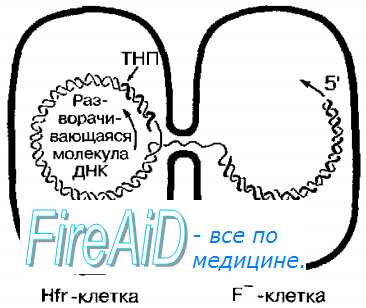 Рис. 4-17. Hfr-конъюгация. Hfr-бактерии функционируют при конъюгации как доноры. Перенос ДНК осуществляется линейно и начинается с удвоения места включения F-фактора (то есть с удвоения точки начала переноса — ТНП).
Рис. 4-17. Hfr-конъюгация. Hfr-бактерии функционируют при конъюгации как доноры. Перенос ДНК осуществляется линейно и начинается с удвоения места включения F-фактора (то есть с удвоения точки начала переноса — ТНП).
• На поверхности реципиентных бактерий должны быть рецепторы пилей, имеющие существенное сродство ( к F-пилям, что позволяет образовать стабильную связь между пилями и рецепторами.
• Для эффективной конъюгации у F-фактора должна быть точка начала репликации, распознаваемая репликативными системами хозяина.
• Эффективность Hfr-конъюгации зависит от величины гомологии ДНК. Перенос негомологичного хромосомного материала донора не приведёт к его интеграции с ДНК реципиента.
Источник



