
Классификация вирусов по способу передачи микробиология
Вирусы классифицируют по типу генетического материала, способам репликации, строению и расположению структурных белков (капсидов), а также наличию или отсутствию оболочки.
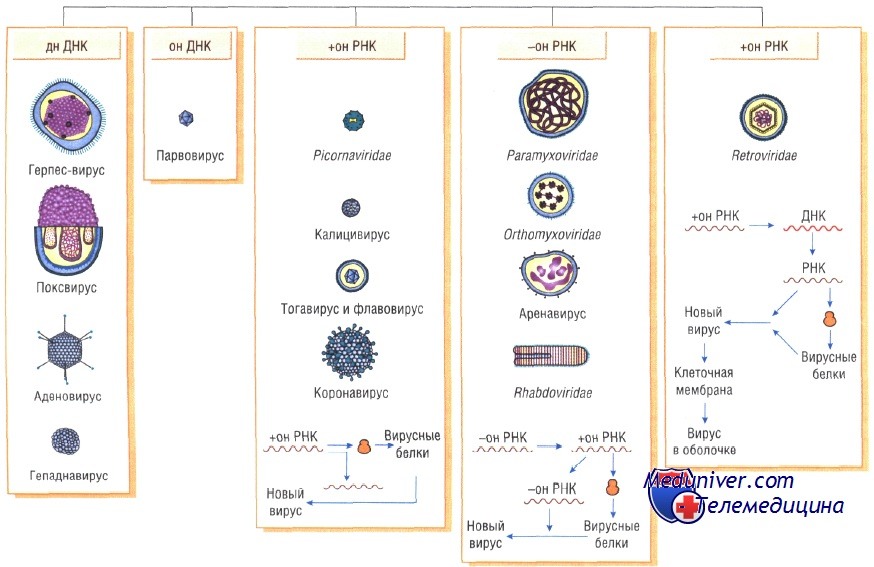
Генетическая структура и способы репликации ДНК-вирусы. Могут быть только двунитевыми и одноните-выми. К. первым относят вирус оспы, герпес-вирусы, аденовирусы, паповавирусы и полиомавирусы. Последние два вируса вызывают развитие доброкачественных (бородавки) и злокачественных (рак шейки матки) опухолей. Вирус гепатита В частично дву- и однонитевой. К однонитевым вирусам относят парвовирусы, вызывающие инфекционную эритему.
Репликация ДНК-вирусов обычно происходит в ядре клеток хозяина и сопровождается продукцией полимераз, воспроизводящих вирусную ДНК. При этом последняя не всегда встраивается в хромосомную ДНК хозяина.
РНК-вирусы. Эти вирусы содержат однонитевую РНК, но различаются по стратегии репродукции, (вирусы, содержащие плюс-однонитевую РНК и минус-однонитевую РНК). У плюс-однонитевых вирусов РНК транслируется в структурные белки и служит матрицей (мРНК) для РНК-зависимой РНК-полимеразы.
В состав минус-однонитевых вирусов входит собственная РНК-зависимая РНК-полимераза, продуцируемая на базе генома вируса мРНК. Последняя в свою очередь может быть матрицей для продукции вирусной (минус-однонитевой) РНК.
Ретровирусы имеют плюс-однонитевую РНК, которая не может выступать в качестве мРНК. Она «переписывается» на ДНК при помощи обратной транскриптазы и встраивается в ДНК хозяина. Последующую транскрипцию с образованием мРНК и вирусной РНК контролирует транскриптаза клеток хозяина.
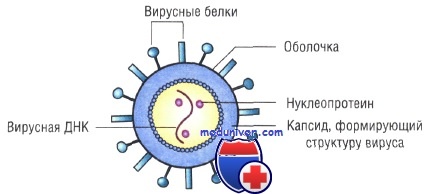
Строение капсидов вирусов. Вирусная нуклеиновая кислота покрыта белковой оболочкой, состоящей из повторяющихся единиц (капсида) с икосаэдрическим (кубическим) или спиральным типами симметрии. Капсиды вирусов с икосаэдрическим типом симметрии имеют практически сферическую форму. Спиральный тип симметрии свойствен РНК-вирусам, капсиды которых окружают нуклеиновую кислоту, располагающуюся в виде спирали.
Капсид состоит из повторяющихся компонентов (капсомеров), количество генов, кодирующих его, снижено, тем самым облегчён процесс сборки вируса.
Оболочка вирусов. В некоторых случаях нуклеиновая кислота и капсидные белки вируса (нуклеокапсид) окружены липидной оболочкой, состоящей из компонентов клетки хозяина или ядерных мембран. Мембрана клетки хозяина изменяется под действием белков, кодируемых вирусом, или гликопротеинов, выступающих в роли рецепторов для других клеток хозяина. Покрытые оболочкой вирусы чувствительны к действию веществ, растворяющих липидную мембрану (например, эфиров).
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Источник
7. Вирусы, их строение, биологические свойства, размножение вирусов, культивирование в живых системах. Классификация вирусов.
Вирусы — мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царству Vira. Явл. облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые к-ты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом.
В основу классификации вирусов положены следующие категории:
тип нуклеиновой кислоты (ДНК или РНК), ее структура, количество нитей;
размер и морфология вирионов, количество капсомеров и тип симметрии;
чувствительность к эфиру и дезоксихолату;
место размножения в клетке;
антигенные свойства и пр.
Морфологию вирусов изучают с помощью электронной микроскопии,т.к. их размеры малы (18-400 нм).
По формам вирионы мб: палочковидные, пулевидные(вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), нитевидной, в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы.
Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид.
Сложные, или оболочечные, вирусы снаружи капсида окружены липопротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок.
Тип симметрии. Капсид может иметь спиральный, кубический или сложный тип симметрии. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида.
Методы культивирования вирусов
Для культивирования вирусов используют культуры клеток, куриные эмбрионы и чувствительных лабораторных животных. Т.к. вирусы е растут на искусственных средах.
Культуры клеток. готовят из тканей животных или человека. Культуры подразделяют на первичные (неперевиваемые), полуперевиваемые и перевиваемые.
Приготовление первичной культуры клеток складывается из нескольких последовательных этапов: измельчения ткани, разъединения клеток путем трипсинизации, отмывания полученной однородной суспензии изолированных клеток от трипсина с последующим суспендированием клеток в питательной среде, обеспечивающей их рост, например в среде 199 с добавлением телячьей сыворотки крови.
Перевиваемые культуры в отличие от первичных адаптированы к условиям, обеспечивающим им постоянное существование invitro, и сохраняются на протяжении нескольких десятков пассажей. Перевиваемые однослойные культуры клеток приготовляют из злокачественных и нормальных линий клеток, обладающих способностью длительно размножатьсяinvitro в определенных условиях. К ним относятся злокачественные клетки HeLa, первоначально выделенные из карциномы шейки матки, Нер-3 (из лимфоидной карциномы), а также нормальные клетки амниона человека, почек обезьяны и др.
К полуперевиваемым культурам относятся диплоидные клетки человека. Они представляют собой клеточную систему, сохраняющую в процессе 50 пассажей (до года) диплоидный набор хромосом, типичный для соматических клеток используемой ткани. Диплоидные клетки человека не претерпевают злокачественного перерождения и этим выгодно отличаются от опухолевых.
О размножении (репродукции) вирусов в культуре клеток судят по цитопатическому действию (ЦПД), которое может быть обнаружено микроскопически и характеризуется морфологическими изменениями кл. Характер ЦПД вирусов используют как для их обнаружения (индикации), так и для ориентировочной идентификации, т. е. определения их видовой принадлежности.
Один из методов индикации вирусов основан на способности поверхности клеток, в которых они репродуцируются, адсорбировать эритроциты — реакция гемадсорбции. Для ее постановки в культуру клеток, зараженных вирусами, добавляют взвесь эритроцитов и после некоторого времени контакта клетки промывают изотоническим раствором хлорида натрия. На поверхности пораженных вирусами клеток остаются прилипшие эритроциты.
Другой метод — реакция гемагглютинации (РГ). Применяется для обнаружения вирусов в культуральной жидкости культуры клеток либо хорионаллантоисной или амниотической жидкости куриного эмбриона.
Количество вирусных частиц определяют методом титрования по ЦПД в культуре клеток. Для этого клетки культуры заражают десятикратным разведением вируса. После 6—7-дневной инкубации их просматривают на наличие ЦПД. За титр вируса принимают наибольшее разведение, которое вызывает ЦПД в 50 % зараженных культур. Титр вируса выражают количеством цитопатических доз.
Более точным количественным методом учета отдельных вирусных частиц является метод бляшек.
Некоторые вирусы можно обнаружить и идентифицировать по включениям, которые они образуют в ядре или цитоплазме зараженных клеток. Куриные эмбрионы. Куриные эмбрионы по сравнению с культурами клеток значительно реже бывают контаминированы вирусами и микоплазмами, а также обладают сравнительно высокой жизнеспособностью и устойчивостью к различным воздействиям. Для получения чистых культур риккетсий, хламидий и ряда вирусов в диагностических целях, а также для приготовления разнообразных препаратов (вакцины, диагностикумы) используют 8—12-дневные куриные эмбрионы. О размножении упомянутых микроорганизмов судят по морфологическим изменениям, выявляемым после вскрытия эмбриона на его оболочках. О репродукции некоторых вирусов, например гриппа, оспы, можно судить по реакции гемагглютинации (РГА) с куриными или другими эритроцитами. К недостаткам данного метода относятся невозможность обнаружения исследуемого микроорганизма без предварительного вскрытия эмбриона, а также наличие в нем большого количества белков и других соединений, затрудняющих последующую очистку риккетсий или вирусов при изготовлении различных препаратов. Лабораторные животные. Видовая чувствительность животных к определенному вирусу и их возраст определяют репродуктивную способность вирусов. Во многих случаях только новорожденные животные чувствительны к тому или иному вирусу (например, мыши-сосунки — к вирусам Коксаки). Преимущество данного метода перед другими состоит в возможности выделения тех вирусов, которые плохо репродуцируются в культуре или эмбрионе. К его недостаткам относятся контаминация организма подопытных животных посторонними вирусами и микоплазмами, а также необходимость последующего заражения культуры клеток для получения чистой линии данного вируса, что удлиняет сроки исследования.
Репродукция вирусов. Стадии взаимодействия вирусов с клеткой.
Размножение вирусов происходит особым, ни с чем не сравнимым способом. Сначала вирионы проникают внутрь клетки, и освобождаются вирусные нуклеиновые кислоты. Затем «заготавливаются» детали будущих вирионов. Размножение заканчивается сборкой новых вирионов и выходом их в окружающую среду.
Встреча вирусов с клетками начинается с его адсорбций, то есть прикрепления к клеточной стенки, плазматической мембране клетки. Причём каждый вирион способен прикрепляться лишь к определённым клеткам, имеющие специальные рецепторы. На одной клетке могут адсорбироваться десятки и даже сотни вирионов. Затем начинается внедрение или проникновение вириона в клетку, которое осуществляет она сама. Этот процесс называется виропексисом.
Клетка как бы «втягивает» прикрепившихся вирионов внутрь. Более просто устроены бактерии не способны сами захватывать вирионы из окружающей среды. Этим, по-видимому, и можно объяснить наличие у поражающих их вирусов сложного и совершенного аппарата, подобно шприцу, впрыскивающего нуклеиновые кислоты.
В зараженной клетке бактериальные ферменты репликации синтезируют комплементарную ей цепь, которая служит матрицей для образования фаговых ДНК. Они соединяются с фаговыми белками, также синтезированные бактериальными ферментами, и новые фаги покидают клетку-хозяина.
Вирусы не способны размножаться на питательных средах — это строгие внутриклеточные паразиты. Более того, в отличие от риккетсий и хламидий, вирусы в клетке хозяина не растут и не размножаются путем деления. Составные части вируса — нуклеиновые кислоты и белковые молекулы синтезируются в клетке хозяина раздельно, в разных частях клетки — в ядре и в цитоплазме. При этом клеточные белоксинтезирующие системы подчиняются вирусному геному, его НК.
Репродукция вируса в клетке происходит в несколько фаз:
Первая фаза — адсорбция вируса на поверхности клетки, чувствительной к данному вирусу.
Вторая фаза — проникновение вируса в клетку хозяина путем виропексиса.
Третья фаза — «раздевание» вирионов, освобождение нуклеиновой кислоты вируса от суперкапсида и капсида.
У ряда вирусов проникновение нуклеиновой кислоты в клетку происходит путем слияния оболочки вириона и клетки-хозяина. В этом случае вторая и третья фазы объединяются в одну.
Источник
4.1. Классификация вирусов
В силу своих особенностей вирусы выделены в отдельное надцарство Vira, в котором по типу нуклеиновой кислоты различают рибовирусы и дезоксирибовирусы (табл. 1).
Подцарства делятся на семейства, которые подразделяются на подсемейства и роды. Вид – совокупность вирусов, имеющих почти идентичные геном (ДНК или РНК), свойства и способность вызывать определенный патологический процесс. Названия семейства имеют окончание viridae, подсемейство – virinae, рода – virus.
Признаки, используемые для классификации вирусов: 1) тип нуклеиновой кислоты – ДНК или РНК; 2) их структура (однонитевая, двунитевая, линейная, кольцевая, фрагментированная, нефрагментированная с повторяющимися и инвертированными последовательностями); 3) структура, размеры, тип симметрии, число капсомеров; 4) наличие или отсутствие внешней оболочки (суперкапсида); 5) антигенная структура; 6) феномены генетических взаимодействий; 7) круг восприимчивых хозяев; 8) географическое распространение; 9) внутриядерная или цитоплазматическая локализация; 10) чувствительность к эфиру и детергентам; 11) путь передачи инфекции.
Для определения принадлежности к семейству ретровирусов обязательно учитывается наличие фермента обратной транскриптазы.
Вирусы, вызывающие инфекционные процессы у человека, входят в состав как ДНК-содержащих, так и РНК-содержащих вирусных семейств (см. табл. 1).
Классификация и некоторые свойства вирусов
Размер вириона, нм
фрагментированная, однонитчатая, кольцевая
Вирусы геморрагических лихорадок
Вирус гепатита Е, калицивирусы
однонитчатая, фрагментированная (-)РНК
Вирусы парагриппа, кори, эпидемического паротита, РС-вирус
Вирусы полиомиелита, Коксаки,
ЕСНО, гепатита А, риновирусы
Вирусы рака, лейкоза, саркомы, ВИЧ
Вирусы лошадиных энцефалитов, краснухи и др.
Вирусы клещевого энцефалита, желтой лихорадки, Денге, японского энцефалита, гепатитов С, G
Вирус бешенства, вирус везикулярного стоматита
Вирусы лихорадки Эбола, Марбург
с однонитчатым участком
Вирус гепатита В
Вирусы простого герпеса, цитомегалии, ветряной оспы, инфекционного мононуклеоза
Вирусы папилломы, полиомы
с замкнутыми концами
Вирус осповакцины, вирус натуральной оспы
4.2. Строение вирусов
По строению различают два типа вирусных частиц – простые и сложные. В составе простых вирионов есть ДНК или РНК и белки. У сложных в суперкапсиде содержатся липиды, полисахариды.
Внутренняя структура простых и сложных вируосв сходна, сердцевина вируса – вирусный геном, который содержит от 3 до 100 и более генов.
Морфология и структура вирусов. Простые вирусы имеют одну белковую оболочку – капсид, который состоит из капсомеров – белковых молекул, форма укладки которых определяет тип симметрии. Капсид представлен -спиральными белками, способными к полимеризации.
Сложные вирусы имеют внешнюю оболочку – суперкапсид, расположенную поверх капсида. В состав суперкапсида входит внутренний белковый слой – М-белок, затем более объемный слой липидов и углеводов, извлеченных из клеточных мембран клетки хозяина. Вирусспецифические гликопротеиды проникают внутрь суперкапсида, образуя фигурные выпячивания (шипы, фибры), которые выполняют рецепторную функцию.
Различают 3 типа симметрии: 1) спиральный, когда капсомеры укладываются по спирали – винтообразная структура нуклеокапсида; 2) кубический (икосаэдрический), когда капсомеры укладываются по граням многогранника (12-20-гранника) – в основе лежит фигура икосаэдра (20-гранника). В зависимости от типа перегруппировки и числа субъединиц число капсомеров будет равным 30, 20, или 12. Вирионы со сложным капсидом, построенным более чем из 60 капсомеров, содержат группы из 5 субъединиц – пентамеры, или из 6 субъединиц – гексамеры; 3) смешанный тип симметрии (у бактериофагов).
Комплекс капсида и генома вируса называют нуклеокапсид. Сложные вирусы имеют суперкапсид (пеплос). Эта поверхностная оболочка вируса, состоит из липидов и белков клеточного происхождения.
Вирусные белки бывают: 1) структурные; 2) неструктурные.
Среди структурных различают: капсидные – входят в состав капсомеров и образуют футляр, защищающий нуклеиновую кислоту; суперкапсидные – это гликопротеиды, которые формируют шипы на поверхности суперкапсида и выполняют: адресную функцию – узнают чувствительную клетку и адсорбируются на ней; прикрепительные белки, которые взаимодействуют со специфическими рецепторами клетки; белки слияния – способствуют слиянию вирусной и клеточной мембран и приводят к образованию симпластов; геномные – обладают антигенными свойсвами, участвуют во взаимодействии с клеткой.
Среди неструктурных белков различают: предшественники вирусных белков (нестабильные); РНК- и ДНК-полимеразы – участвуют в репликации вирусного генома; регуляторные белки – участвуют в репродукции вируса.
Функции белков: обладают антигенными и иммуногенными свойствами; участвуют в распознавании клетки и взаимодействии с ней; защищают геном от нуклеаз; обеспечивают тип симметрии.
Липиды входят в состав суперкапсида и представляют смесь нейтральных фосфо- и гликолипидов, многие из них – продукты мембраны клеток хозяина.
Они обусловливают инфекционность, чувствительность или устойчивость к эфиру; стабилизируют вирусную частицу.
Углеводы входят в состав гликопротеидов суперкапсида. Углеводы и липиды – составная часть гемагглютинина, который вызывает склеивание эритроцитов и обладает антигенной специфичностью.
Различают вирионные и вирусиндуцированные ферменты вирусов. К вирионным относят ферменты транскрипции и репликации (ДНК и РНК-полимеразы); обратную транскриптазу (у ретровирусов), АТФ-азы, эндо- и экзонуклеазы, нейраминидазы.
К вирусиндуцированным относятся ферменты, о которых имеется только информация в вирусном геноме, а появляются они в клетке. Это РНК-полимеразы тога-, орто-, пикорна- и парамиксовирусов; и ДНК-полимеразы у покс- и герпесвирусов.
Нуклеиновые кислоты обеспечивают наследственные признаки; являются хранителями генетической информации; необходимы для репродукции вирусов, многие из них могут вызывать инфекционный процесс самостоятельно, достаточно их проникновения в клетку.
Вирусная ДНК. Молекулярная масса равна 1,10 6 -1,10 8 дальтон. ДНК может быть одно- или двунитчатой, фрагментированной и сверхспирализованной, линейной или кольцевой, содержит несколько сотен генов. В каждой нити ДНК есть нуклеотидные последовательности, а на концах есть прямые или инвертированные (повернутые на 180 о ) повторы, которые являются маркерами для отличия вирусной ДНК от клеточной. Эти повторы обеспечивают способность ДНК замыкаться в кольцо для последующих репликации, транскрибирования и встраивания в клеточный геном. Генетическая информация инфекционной ДНК транслируется на мРНК в клетке с помощью полимераз.
Вирусная РНК может быть одно- и двунитчатой, линейной, кольцевой, фрагментированной. У РНК-содержащих вирусов генетическая информация закодирована в РНК таким же кодом, как в ДНК всех других вирусов и клеточных организмов. Вирусные РНК по своему химическому составу не отличаются от РНК клеточного происхождения, но характеризуются разной структурой.
Наряду с типичной для всех РНК однонитевой формой у ряда вирусов имеется двунитевая РНК. В составе однонитевых РНК имеются спиральные участки типа двойной спирали ДНК, образующиеся вследствие спаривания комлементарных азотистых оснований. Вирусы с однонитчатой РНК делятся на 2 группы: (+)РНК (положительный геном) и (-)РНК (отрицательный геном). Вирусная (+)РНК инфекционная и обладает функциями информационной РНК. Она может передовать генетическую информацию на рибосомы, как иРНК. Вирусы с отрицательным геномом не обладают инфекциозностью, т.к. нить (-)РНК выполняет только наследственную функцию и не обладает функцией иРНК. В зараженной клетке на матрице вирусной геномной РНК с помощью фермента транскриптазы осуществляется синтез РНК-комплементарной геному.
Нити (+)РНК вирусов в отличие от (-)РНК имеют специальные концы в виде «шапочки» для специфического узнавания рибосом.
Патогенность вирусов обусловлена совокупностью их свойств: способностью проникать в макроорганизм, связываться с клеточными мембранами и проникать в клетку, управлять метаболизмом и белоксинтезирующей функцией клетки, обеспечивать транскрипцию и репликацию собственного генома и осуществлять весь цикл репродукции вирусов. Все эти свойства зависят от генома вирусов и наличия соответствующих структурных белков и ферментов. Репродукция вирусов приводит к развитию патологии: цитопатогенному (разрушающему) действию, развитию воспаления, повреждению различных клеток и тканей.
4.3. Взаимодействие вируса с клеткой
Различают 3 типа взаимодействия: продуктивный тип – репродукция завершается образованием вирусного потомства; абортивный тип – не образуются новые вирусные частицы, т.к. инфекционный процесс прерывается на одном из этапов; интегрированный тип – вирогения – характеризуется встраиванием вирусной ДНК в хромосому клетки хозяина.
РНК-вирусы размножаются в цитоплазме, кроме вирусов гриппа и ретровирусов, которые осуществляют это в ядре. Однако ДHК-поксвирусы репродуцируются в цитоплазме.
Репродукция вирусов протекает в несколько стадий (рис. 7).
1. Адсорбция вируса на специфических рецепторах чувствительной клетки благодаря белкам прикрепления (адгезинам) и адресным. Число специфических рецепторов на поверхности одной клетки 10 4 -10 5 . Белки адгезины имеют форму нитей (фибры у аденовирусов) или шипов у орто-, парамиксовирусов, рабдовирусов. Адсорбция определяется неспецифическими слоями межмолекулярного натяжения и специфической комплементарностью рецепторов чувствительных клеток и вирусов. Вначале происходит единичная связь вириона с рецептором – такое прикрепление непрочное – адсорбция носит обратимый характер. Чтобы наступила необратимая адсорбция должны появиться множественные связи между рецептором вируса и рецептором клетки, т.е. стабильное мультивалентное прикрепление.
На клетках существуют различные структуры-рецепторы, к которым прикрепляются вирусы своими рецепторами. У орто- и парамиксовирусов их роль выполняют ганглиозиды (сиалосодержащие гликолипиды), у вируса иммунодефицита человека (ВИЧ) – гликопротеид 120 и др. Примеры клеточных рецепторов: CD4 – молекула для ВИЧ, рецепторы для С3-комплемента – для вируса Эпштейна-Барра, b-адренергические рецепторы – для реовирусов.
2. Проникновение вируса в клетку может идти двумя путями: виропексиса и слияния вирусной и клеточной мембран.
При виропексисе (эндоцитозе) происходит инвагинация участка клеточной мембраны, образование внутриклеточной вакуоли, а далее вакуоль с вирусом может попадать в разные участки цитоплазмы или в ядро клетки.
Процесс слияния осуществляется с помощью вирусных белков капсидной или суперкапсидной оболочек, которые сливаются с плазматической мембраной клетки хозяина.
У парамиксовирусов имеется специальный F-белок, вызывающий слияние клеточных и вирусных мембран. Сходные белки имеются у других вирусов. У вируса гриппа это гемагглютин, который обусловливает адсорбцию его на мембране клетки. Белки слияния служат важнейшими факторами вирулентности вирусов. Они приводят к образованию клеточного синцития (например, при ВИЧ-инфекции).
3. «Раздевание» вирионов или депротеинизация – это процесс освобождения нуклеиновой кислоты вируса от окружающей ее оболочки с последующим проникновением ее в цитоплазму или в ядро клетки. «Раздевание» вириона начинается сразу же после его прикрепления к клеточным рецепторам и продолжается в эндоцитарной вакуоли, а также в ядерных порах и околоядерном пространстве.
4. Биосинтез компонентов вирусов. Нуклеиновая кислота, проникшая в клетку, несет генетическую информацию, которая конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет метаболизм клетки и заставляет ее синтезировать вирусные белки и нуклеиновые кислоты, которые идут на построение вирусного потомства.
Так как генетический аппарат вирусов различен, то передача наследственной информации и синтез ДНК и РНК отличаются.
При инфицировании ДНК-содержащим вирусом идет транскрипция ДНК-вируса на иРНК с помощью ДНК-зависимой РНК полимеразы, которая может быть вирусной при репродукции его в цитоплазме или клеточной, если это происходит в ядре (аденовирусы и др.). Причем, если это происходит в цитоплазме (поксвирусы), то вирусная РНК-полимераза считывает часть ДНК-генома и запускает синтез иРНК, а она – образование первичных ферментов для репликации вирусной ДНК. Эти ферменты индуцируют считывание второй части исходной ДНК – появляется «поздняя» иРНК, обеспечивающая синтез структурных белков.
При инфицировании РНК-содержащим вирусом РНК синтезируется с помощью РНК-полимеразы на матрице вирусной РНК; синтез вирусных белков происходит в цитоплазме, а РНК в ядре или в цитоплазме (пикорнавирусы, тогавирусы).
Для (+)РНК-нитевых вирусов (флави-, пикорна-, тогавирусы) функцию информационной РНК выполняет сам геном, который является матрицей для новых молекул РНК, на основе которых в рибосомах синтезируются вирусные белки.
У (-)РНК-вирусов (орто-, парамиксо-, рабдовирусы) геном не выполняет функцию информационной РНК, не обладает инфекционностью, но вирусы имеют РНК-полимеразы, необходимые для синтеза РНК, комлементарных геному, т.е. мРНК, которые обеспечивают синтез вирусных белков.
Иначе осуществляется репликация РНК-содержащих ретровирусов (онкогенные, ВИЧ), в составе которых есть обратная транскриптаза или ревертаза (см. рис. 7). Уникальность этого фермента состоит в его способности индуцировать синтез цепи вирусной ДНК на матрице вирусной РНК. Этот процесс называется обратной транскрипцией. На матрице одной ДНК-цепи синтезируется комплементарная ей вторая; образовавшаяся двунитевая ДНК переносится в ядро. Клеточная ДНК подвергается сплайсингу (под влиянием эндонуклеаз) с образованием рекомбинантов с этой вирусной ДНК. Возникает ДНК-провирус. С помощью клеточной ДНК-зависимой РНК-полимеразы интегрированный в ДНК клетки ДНК-провирус считывается с последующим синтезом вирусных (+)РНК и мРНК, которые определяют образование вирусных структурных белков и ферментов. Продолжающийся синтез цепей ДНК обеспечивает новые вирионы геномом.
Количество генов в вирусном геноме ограничено, поэтому есть дополнительные механизмы для передачи большей информации, чем несет вирусная нуклеиновая кислота. Например, транскрипция информации с переписывающихся участков ДНК на иРНК может происходить путем сплайсинга (вырезание бессмысленных кодонов и сшивание концов), а также вследствие считывания антикодонами тРНК с разных нуклеотидов одной и той же молекулы иРНК, в результате возникают новые триплеты и увеличивается транслируемая информация.
Формирование нуклекапсидов происходит, тогда, когда синтезированные вирусные нуклеиновые кислоты и белки специфично узнают друг друга и соединяются гидрофобными солевыми и водородными связями. Основой самосборки простых вирионов служит способность вирусных полипептидов соединяться в капсомеры, образующие многогранник. Полипептиды могут также окружать в виде спирали вирусную нуклеиновую кислоту.
Для вирусов важен синтез М-белка (матриксный белок), который участвует в сборке вирионов.
Нередко простые вирионы монтируются на репликативных комплексах, которыми служат мембраны эндоплазматического ретикулума. Сборка нуклеокапсида сложных вирионов начинается на репликативных комплексах и продолжается на плазматической мембране, где присутствуют гликопротеиды суперкапсида.
Нуклеокапсиды вируса герпеса и многих ДНК-содержащих вирусов монтируются в ядре клетки на ее мембране. Затем они отпочковываются и приобретают суперкапсидную оболочку. Окончательное формирование вириона осуществляется в мембранах эндоплазматического ретикулума и в аппарате Гольджи.
Выход вирусов из клетки происходит: 1) путем «взрыва» оболочки (клетка при этом погибает), что характерно для вирусов, не имеющих суперкапсида (пикорнавирусы); 2) путем почкования, что присуще вирусам, имеющим суперкапсид. На заключительном этапе сборки нуклеокапсиды фиксируются на клеточной плазматической мембране и выпячивают ее, образуется «почка», которая затем отделяется от клетки (орто-, парамиксо-, рабдовирусы). Клетка при этом сохраняет жизнеспособность.
Время, необходимое для репродукции, колеблется от 5-6 часов (пикорновирусы) до нескольких суток (вирусы кори, аденовирусы).
Интеграция с клеточным геномом. Нуклеиновые кислоты (ДНК и РНК) вирусов могут быть включены в геном клетки с помощью эндонуклеаз, рестриктаз и лигаз. Вирусная ДНК в кольцевой форме интегрируется в клеточный геном. Место включения в геном определяется гомологичными нуклеотидными последовательностями определенных участков (ДНК-сайтов). После включения в геном клетки ДНК-вируса становится провирусом, который может изменять клеточный метаболизм, что приводит к возникновению аутоиммунных, хронических заболеваний и опухолей. Под влиянием физических и химических факторов ДНК-провирус вырезается из клеточной хромосомы и становится обычным вирусом.
Геном РНК-вирусов включается в клеточный геном при участии обратной транскриптазы, образующей ДНК-транскрипт одной цепи на матрице РНК (см. рис. 7). Образовавшаяся нить ДНК служит матрицей для образования второй нити, а затем двуцепочечный ДНК-транскрипт замыкается в кольцо и встраивается в клеточный геном. Этот процесс интеграции РНК-вируса в геном клетки называют вирогенией. Как и в случае с ДНК-провирусами в клетке могут изменяться свойства, что приводит к заболеваниям.
Виды вирусной инфекции. На клеточном уровне выделяют автономные инфекции, если вирусный геном реплицируетсЯ независимо от клеточного и интеграционные инфекции, если вирусный геном включается в состав клеточного. Автономная инфекция делится на продуктивную, при которой образуется инфекционное потомство и абртивную, при которой инфекционный процесс обрывается и новые инфекционные частицы не образуются.
По течению различают острую и персистирующую инфекции. Острая инфекция в зависимости от судьбы зараженной клетки делится на цитолитическую и нецитолитическую, цитолитическая заканчивается образованием ЦПД. Она сопровождает продуктивную фазу взаимодействия вируса с клеткой. Персистирующая инфекция может протекать в виде вирусоносительства, латентной, хронической или медленной инфекции. Латентная – это инфекция без выделения вируса в окружающую среду. Хроническая инфекция сопровождается выделением вируса и периодами ремиссии и обострения. Медленная инфекция характеризуется длительным инкубационным периодом и последующим длительным прогрессирующим течением с летальным исходом.
На уровне организма вирусные инфекции делятся на очаговые – когда размножение вирусов происходит у входных ворот и генерализованные – когда вирус разносится по различным органам и тканям.
Особенности вирусных инфекций.
Внутриклеточный паразитизм вирусов приводит к гибели клеток.
Размножение некоторых вирусов в клетках иммунной системы приводит к развитию иммунодефицитных состояний (вирус кори, ВИЧ, гепатита В, С).
Некоторые вирусы способны интегрировать с геномом клетки хозяина (ВИЧ, онкогенные РНК-содержащие вирусы).
Некоторые вирусы обладают тератогенным действием (вирус цитомегалии, краснухи).
Диагностика вирусных инфекций не проводится в каждом случае заболевания из-за массовости.
Вирусы могут вызывать медленные инфекции (вирусы кори, ВИЧ, бешенства, гепатит В, герпес).
Некоторые вирусы могут провоцировать развитие опухолей (герпесвирусы, гепатита В, С, аденовирусы). гепатита В, С
Тут вы можете оставить комментарий к выбранному абзацу или сообщить об ошибке.
Источник



