Классификация синапсов по способу передачи сигнала
Синaпс – специализированный контакт между нервными клетками (или нервными и другими возбудимыми клетками), обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов нервные клетки объединяются в нервные сети, которые осуществляют обработку информации. Взаимосвязь между нервной системой и периферическими органами и тканями также осуществляется при помощи синапсов.
Классификация синапсов
По морфологическому принципу синапсы подразделяют на:
нейро-мышечные (аксон нейрона контактирует с мышечной клеткой);
нейро-секреторные (аксон нейрона контактирует с секреторной клеткой);
нейро-нейрональные (аксон нейрона контактирует с другим нейроном):
аксо-соматические (с телом другого нейрона), аксо-аксональные (с аксоном другого нейрона), аксо-дендритические (с дендритом другого нейрон).
По способу передачи возбуждения синапсы подразделяют на:
электрические (возбуждение передается при помощи электрического тока);
химические (возбуждение передается при помощи химического вещества):
адренергические (возбуждение передается при помощи норадреналина), холинергические (возбуждение передается при помощи ацетилхолина), пептидергические, NO -ергические, пуринергические и т. п.
По физиологическому эффекту синапсы подразделяют на:
возбуждающие (деполяризуют постсинаптическую мембрану и вызывают возбуждение постсинаптической клетки);
тормозные (гиперполяризуют постсинаптическую мембрану и вызывают торможение постсинаптической клетки).
Ультраструктура синапсов
Все синапсы имеют общий план строения (рис. 1).
Конечная часть аксона (синаптическое окончание), подходя к иннервируемой клетке, теряет миелиновую оболочку и образует на конце небольшое утолщение (синаптическую бляшку). Ту часть мембраны аксона, которая контактирует с иннервируемой клеткой, называют пресинаптической мембраной. Синаптическая щель – узкое пространство между пресинаптической мембраной и мембраной иннервируемой клетки, которое является непосредственным продолжением межклеточного пространства. Постсинаптическая мембрана – участок мембраны иннервируемой клетки, контактирующий с пресинаптической мембраной через синаптическую щель.
Рис. 1. Ультраструктура химического и электрического синапса.
Особенности ультраструктуры электрического синапса (см. рис. 1):
узкая (около 5 нм) синаптическая щель; наличие поперечных канальцев, соединяющих пресинаптическую и постсинаптическую мембрану.
Особенности ультраструктуры химического синапса (см. рис. 1):
широкая (20–50 нм) синаптическая щель; наличие в синаптической бляшке синаптических пузырьков (везикул), заполненных химическим веществом, при помощи которого передается возбуждение; в постсинаптической мембране имеются многочисленные хемочувствительные каналы (в возбуждающем синапсе – для Nа+ , в тормозном – для Cl – и К +), но отсутствуют потенциалчувствительные каналы.
Механизм передачи возбужденияв электрическом синапсе
Механизм проведения возбуждения аналогичен механизму проведения возбуждения в нервном волокне. Во время развития ПД происходит реверсия заряда пресинаптической мембраны. Электрический ток, возникающий между пресинаптической и постсинаптической мембраной, раздражает постсинаптическую мембрану и вызывает генерацию в ней ПД (рис. 2).
Рис. 2. Передача возбуждения в электрическом синапсе.
Этапы и механизмы передачи возбуждения в возбуждающем химическом синапсе
Передача возбуждения в химическом синапсе – сложный физиологический процесс, протекающий в несколько этапов. На пресинаптической мембране осуществляется трансформация электрического сигнала в химический, который на постсинаптической мембране снова трансформируется в электрический сигнал.
Синтез медиатора
Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Ферменты, необходимые для синтеза медиатора, образуются в теле нейрона и доставляются в синаптическое окончание путем медленного (1–3 мм/сут) аксонного транспорта. Другие медиаторы (пептиды и др.) синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого (400 мм/сут) аксонного транспорта. Синтез медиатора и образование синаптических пузырьков осуществляется непрерывно.
Секреция медиатора
Содержимое синаптических пузырьков может выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10000 молекул.
Для активации экзоцитоза необходимы ионы Са++ . В состоянии покоя уровень Са++ в синаптическом окончании низок и выделения медиатора практически не происходит. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической мембраны и открытию потенциалчувствительных Са++ -каналов. Ионы Са++ поступают в цитоплазму синаптического окончания (рис. 3, А,Б) и активируют опорожнение синаптических пузырьков в синаптическую щель (рис. 3, В).
Рис. 3. Передача сигнала в возбуждающем химическом синапсе. А — Д – последовательность процессов при срабатывании химического синапса; Е – деполяризация постсинаптической мембраны (ВПСП).
Взаимодействие медиатора с рецепторами постсинаптической мембраны
Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных Na+ -каналов (рис. 3, Г). Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через которые в клетку входят ионы Na+ (рис. 3, Д). В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП) (рис. 3, Е).
Инактивация медиатора
Ферменты, находящиеся в синаптической щели, разрушают молекулы медиатора. В результате происходит закрытие Na+ -каналов и восстановление МП постсинаптической клетки. Некоторые медиаторы (например, адреналин) не разрушаются ферментами, а удаляются из синаптической щели путем быстрого обратного всасывания (пиноцитоза) в синаптическое окончание.
Генерация ПД
В нейро-мышечном синапсе амплитуда единичного ВПСП достаточно велика. Поэтому для генерации ПД в мышечной клетке достаточно прихода одного нервного импульса. Генерация ПД в мышечной клетке происходит в области, окружающей постсинаптическую мембрану.
В нейро-нейрональном синапсе амплитуда ВПСП значительно меньше и недостаточна для того, чтобы деполяризовать мембрану нейрона до КУД. Поэтому для генерации ПД в нервной клетке требуется возникновение нескольких ВПСП. ВПСП, образовавшиеся в результате срабатывания разных синапсов, электротонически распространяются по мембране клетки, суммируются и генерируют образование ПД в области аксонного холмика. Мембрана нейрона в области аксонного холмика обладает низким электрическим сопротивлением и имеет большое количество потенциалчувствительных Na+ -каналов.
Особенности работы тормозного химического синапса
В тормозном химическом синапсе молекулы медиатора, взаимодействуя с рецепторами постсинаптической мембраны, вызывают открытие К+ — и Cl – -хемочувствительных каналов. Вход в клетку Cl– и дополнительная утечка из клетки К+ приводят к гиперполяризации постсинаптической мембраны, которую называют тормозным постсинаптическим потенциалом (ТПСП) . Возникшая гиперполяризация, во-первых, снижает возбудимость клетки. Во-вторых, ТПСП может нейтрализовать возникший в другом месте клетки ВПСП.
Свойства синапсов
Сравнительная характеристика свойств электрических и химических синапсов приведена в табл. 1.
Одностороннее проведение возбуждения в химическом синапсе связано с его функциональной асимметрией: молекулы медиатора выделяются только на пресинаптической мембране, а рецепторы медиатора расположены только на постсинаптической мембране.
Высокая утомляемость химического синапса объясняется истощением запасов медиатора. Утомляемость электрического синапса соответствует утомляемости нервного волокна.
Низкая лабильность химического синапса определяется главным образом периодом рефрактерности хемочувствительных каналов на постсинаптической мембране.
Синаптическая задержка – время от момента возникновения возбуждения в пресинаптической мембране до момента возникновения возбуждения в постсинаптической мембране. Относительно длительное время синаптической задержки в химическом синапсе (0,2–0,7 мс) затрачивается на вход Са++ в синаптическое окончание, экзоцитоз, диффузию медиатора.
Чувствительность синапса к внешним воздействиям определяется характером процессов, протекающих в синапсе при передаче возбуждения. Химические синапсы чувствительны к действию химических веществ, влияющих на синтез и секрецию медиатора, взаимодействие медиатора с рецептором.
Таблица 1.Свойства электрических и химических синапсов
Источник
Физиология человека и животных
Разделы
6. Основы межклеточного взаимодействия в организме. Физиология синапсов
Переход возбуждения от нейрона к нейрону или к мышечной или железистой клетке происходит в местах особых контактов – синапсов (Термин «синапс» ввёл Ч. Шеррингтон в 1897 г.). Синапс – это место контакта двух возбудимых клеток (в частности, нервного окончания и мышцы, нервного окончания одного нейрона и тела другого (вставочного) нейрона, нервного окончания и железистой клетки), обеспечивающее передачу нервных импульсов с одной возбудимой клетки на другую. В своем составе синапс имеет пре- и постсинаптическую мембраны, между которыми находится синаптическая щель. Синапсы могут быть аксодендритные (между окончанием аксона и дендритом следующего нейрона), аксосоматические, реже – сома-соматические, дендродендритные и дендросоматические.
Классификация синапсов:
1. По виду соединяемых клеток:
— межнейронные синапсы – находятся в ЦНС и вегетативных ганглиях;
— нервно-мышечные синапсы — соединяют аксоны мотонейрона с мышечным волокном.
— возбуждающие, т.е. запускающие генерацию ПД;
— тормозные, т.е. препятствующие возникновению ПД.
3. По способу передачи сигнала:
— химические синапсы – передача осуществляется с помощью химического
— электрические синапсы – ПД непосредственно (электротонически) переда-
ется на постсинаптическую клетку;
— смешанные синапсы – наряду с химической передачей имеются участки с
электротоническим механизмом передачи (например, в реснитчатом
ганглии птиц, спинном мозге лягушки).
4. По природе нейромедиатора:
— холинергические (медиатор – ацетилхолин);
6. По местоположению:
— центральные (головной и спинной мозг);
В электрических синапсах возбуждение передается электротонически, за счет локальных круговых токов между пре- и постсинаптической мембранами. Ширина синаптической щели в них составляет 1-2 нм, между синаптическими мембранами существуют узкие щелевые контакты, обладающие низким электрическим сопротивлением. В них почти нет утечки через внеклеточную среду, поэтому изменения потенциала в пресинаптической мембране могут эффективно передаваться на постсинаптическую мембрану, в которой под действием потенциалов действия с пресинаптической мембраны меняется ионная проницаемость и генерируются свои потенциалы действия.
Электрические синапсы встречаются чаще у беспозвоночных и низших позвоночных (например, у ракообразных). В стволе мозга млекопитающих электрические синапсы имеются в ядрах тройничного нерва и в нижней оливе мозгового ствола.
В электрических синапсах проведение возбуждения происходит очень быстро, практически без синаптической задержки, ток возможен в обоих направлениях (но легче – в одном, от пресинаптического окончания к постсинаптической мембране). Электрические синапсы дают возможность получать постоянные, повторяющиеся реакции и синхронизировать активность многих нейронов.
Источник
Классификация синапсов по способу передачи сигнала
Механизм химической передачи нервных импульсов через синапс сводится к тому, что содержимое синаптических пузырьков (медиатор) поступает в виде небольших порций (квантов) в синаптическую щель и взаимодействует затем с рецепторными белками постсинаптической мембраны. Это вызывает деполяризацию мембраны и возбуждение следующего нейрона. Ультраструктурные особенности синапса и механизм передачи импульсов определяют строгую однонаправленность передачи импульсов, что лежит в основе проведения импульсов по рефлекторным дугам.
В зависимости от того, какое вещество выполняет роль нейромедиатора, синапсы подразделяются на холинергические (медиатор — ацетилхолин), адренергические (адреналин и норадреналин), дофаминергические (дофамин), серотонинергические (серотонин), пептидергические (медиаторы — пептиды и аминокислоты, например, мет-энкефалин, гамма-аминомасляная кислота, глицин и др.).
Нейрохимические синапсы подразделяются в функциональном отношении на две противоположные по своему значению группы — возбуждающие и тормозные. Свойства этих синапсов зависят как от медиаторов, так и от ультраструктурных особенностей синапсов. Так, некоторые медиаторы (например, глутамат) характерны для возбуждающих синапсов, а в тормозных синапсах медиатором является гамма-аминомасляная кислота. Предполагают, что в возбуждающих синапсах к постсинаптической мембране прилежит электронно-плотное вещество, в связи с чем синапс приобретает асимметричное строение.
В пресинаптических частях таких соединений содержатся круглые синаптические пузырьки. Тормозные синапсы имеют симметричное строение. Синаптическая щель у них сужена и в пресинаптической части содержатся уплощенные синаптические пузырьки.
Кроме нейрохимических синапсов между нервными клетками (преимущественно между дендритами или телами нейронов) возникают электротонические синапсы. Последние у млекопитающих встречаются редко и по строению соответствуют щелевым контактам. Они проводят возбуждение благодаря формированию трансмембранных каналов — коннексонов.
Каждый нейрон на своей поверхности имеет огромное количество (до 10000) синапсов. Интеграция импульсов в пределах отдельного нейрона происходит так: от синапсов, расположенных на дендритах и теле, по его плазмолемме передается импульс к аксонному холмику (генераторному пункту нейрона), где путем суммирования всех возбуждающих и тормозных импульсов возникает результирующий потенциал действия.
Синаптические структуры обладают высокой чувствительностью к действию токсических факторов, в том числе психотропных отравляющих веществ. Их изменения играют важную роль в механизмах реактивности нервных тканей.
Рецепторные нервные окончания подразделяются на две группы: экстерорецепторы, воспринимающие раздражения из внешней среды, и интерорецепторы, служащие для восприятия раздражений из внутренней среды организма. В зависимости от специфичности раздражения, воспринимаемого рецептором, различают следующие чувствительные нервные окончания: механорецепторы, барорецепторы, хеморецепторы, терморецепторы, болевые рецепторы (ноцицепторы). Все рецепторы по особенностям их строения подразделяют на свободные и несвободные нервные окончания.
Первые из них состоят только из конечных разветвлений периферического отростка чувствительного нейрона; вторые — имеют в составе рецептора кроме нервного отростка глиальный компонент, а нередко и соединительнотканную капсулу. В последнем случае несвободные рецепторные окончания называют инкапсулированными. К числу таких рецепторов относятся пластинчатые, осязательные тельца, концевые колбы, мышечные веретена и др.
Эффекторные нервные окончания подразделяются на двигательные и секреторные. Двигательный, или моторный, эффектор передает нервные импульсы на рабочие органы и ткани. В скелетных мышцах они называются нервно-мышечными (аксомышечными) окончаниями. Терминальное разветвление аксона двигательного нейрона заканчивается булавовидным расширением. Оно соответствует пресинаптической части нервно-мышечного синапса, содержит скопление синаптических пузырьков и митохондрий, ограничено пресинаптической мембраной характерного строения.
Плазмолемма мышечного волокна в этой области образует многочисленные складки и углубления. В постсинаптической части волокна находится зернистая саркоплазма с большим количеством митохондрий и овальных ядер. Синаптическая щель имеет ширину 50-100 нм. У человека медиатором в нервно-мышечных синапсах является ацетилхолин. Моторные окончания в гладкой мышечной ткани, а также секреторные эффекторы имеют вид тонких пучков аксонов или их одиночных терминалей, прилежащих к клеткам иннервируемых тканей.
Возрастные изменения нервных тканей связаны с постепенным уменьшением запаса нервных клеток, особенно — чувствительных нейронов, а также снижением уровня метаболических процессов, что выражается в закономерном накоплении включений липофусцина («пигмент изнашивания») в нейроплазме.






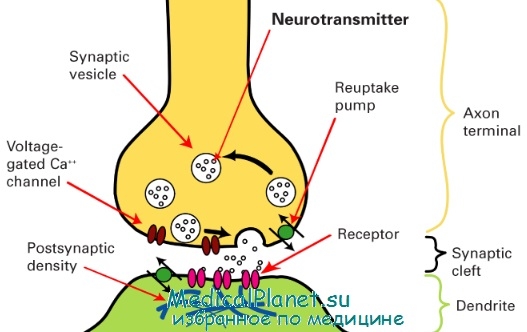 Схема строения синапса
Схема строения синапса 


