
Биология. 11 класс
§ 16. Клеточный цикл. Репликация ДНК
Клеточный цикл . Все новые клетки образуются путем деления уже существующих. Этот принцип, как уже отмечалось, сформулировал Р. Вирхов еще в середине XIX в. Деление клеток обеспечивает непрерывность существования жизни на нашей планете. Именно благодаря ему осуществляются различные способы бесполого и полового размножения организмов. В основе процессов роста, регенерации и индивидуального развития многоклеточных организмов также лежит деление клеток.
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
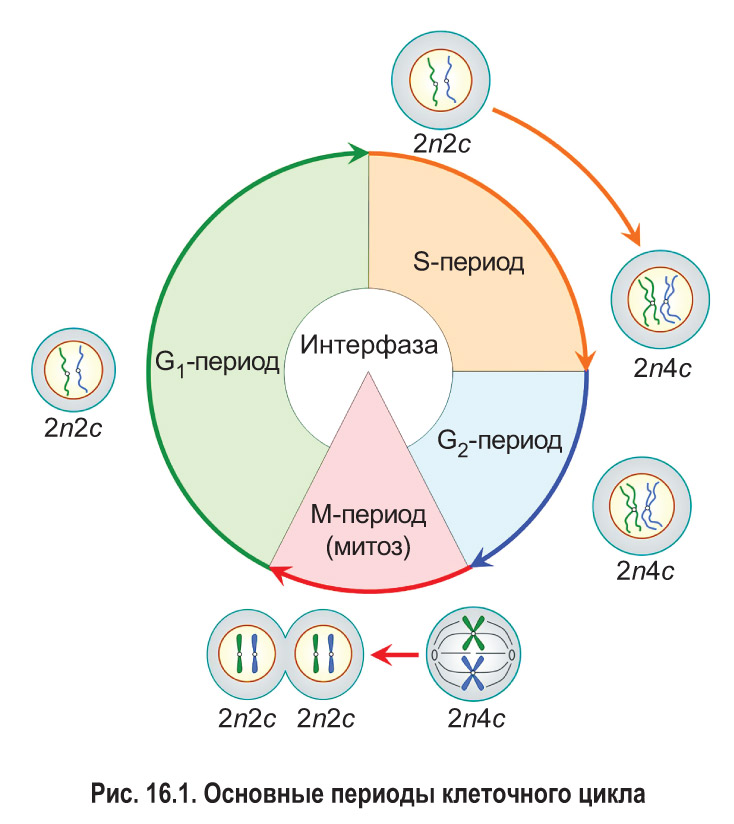 Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Итак, интерфаза обычно включает три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На протяжении всей интерфазы хромосомы не спирализованы. Они располагаются в ядре клетки в виде хроматина .
После завершения интерфазы начинается деление клетки. Основным способом деления клеток эукариот является митоз, поэтому данный этап клеточного цикла обозначают как М-период. При митозе происходит спирализация хроматина . Это приводит к формированию компактных двухроматидных хромосом. После этого сестринские хроматиды каждой хромосомы отделяются друг от друга и затем попадают в разные дочерние клетки. Значит, дочерние клетки, образовавшиеся в результате митоза и вступающие в новый клеточный цикл, имеют набор 2n2c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Источник
Репликация ДНК
Репликация — это механизм самокопирования и основное свойство наследственного материала, которым выступают молекулы ДНК.
Особенностью ДНК является то, что обычно ее молекулы состоит из двух комплементарных друг другу цепей, образующих двойную спираль. В процессе репликации цепи материнской молекулы ДНК расходятся, и на каждой строится новая комплементарная цепь. В результате из одной двойной спирали образуется две, идентичные исходной. Т. е. из одной молекулы ДНК образуются две, идентичные матричной и между собой.
Таким образом, репликация ДНК происходит полуконсервативным способом, когда каждая дочерняя молекула содержит одну материнскую цепь и одну вновь синтезированную.
У эукариот репликация происходит в S-фазе интерфазы клеточного цикла.
Описанный ниже механизм и основные ферменты характерны для подавляющего большинства организмов. Однако бывают исключения, в основном среди бактерий и вирусов.
Расхождение цепей исходной молекулы ДНК обеспечивает фермент геликаза, или хеликаза, который в определенных местах хромосом разрывает водородные связи между азотистыми основаниями ДНК. Хеликазы перемещаются по ДНК с затратой энергии АТФ.
Чтобы цепочки снова не соединились, они удерживаются на расстоянии друг от друга дестабилизирующими белками. Белки выстраиваются в ряд со стороны пентозо-фосфатного остова цепи. В результате образуются зоны репликации, называемые репликационными вилками.
Репликационные вилки образуются не в любых местах ДНК, а только в точках начала репликации, состоящих из определенной последовательности нуклеотидов (около 300 штук). Такие места распознаются специальными белками, после чего образуется так называемый репликационный глаз, в котором расходятся две цепи ДНК.
Из точки начала репликация может идти как в одном, так и в двух направлениях по длине хромосомы. В последнем случае цепи ДНК расходятся вперед и назад, и из одного репликационного глазка образуются две репликационные вилки.
Репликон — единица репликации ДНК, от точки ее начала и до точки ее окончания.
Поскольку в ДНК цепи спирально закручены относительно друг друга, то разделение их хеликазой вызывает появление дополнительных витков перед репликационной вилкой. Чтобы снять напряжение, молекула ДНК должна была бы проворачиваться вокруг своей оси один раз на каждые 10 пар разошедшихся нуклеодидов, именно столько образуют один виток спирали. В таком случае ДНК бы быстро вращалась с затратой энергии. Но этого не происходит, т. к. природа нашла более эффективный способ справится с возникающим при репликации напряжением спирали.
Фермент топоизомераза разрывает одну из цепей ДНК. Отсоединенный участок проворачивается на 360° вокруг второй целой цепи и снова соединяется со своей цепью. Этим снимается напряжение, т. е. устраняются супервитки.
Каждая отдельная цепь ДНК старой молекулы используется в качестве матрицы для синтеза новой комплементарной себе цепи. Добавление нуклеотидов к растущей дочерней цепи обеспечивает фермент ДНК-полимераза. Существует несколько разновидностей полимераз.
В репликационной вилке к освободившимся водородным связям цепей согласно принципу комплиментарности присоединяются свободные нуклеотиды, находящиеся в нуклеоплазме. Присоединяющиеся нуклеотиды представляют собой дезоксирибонуклеозидтрифосфаты (дНТФ), а конкретно дАТФ, дГТФ, дЦТФ, дТТФ.
После образования водородных связей фермент ДНК-полимераза связывает нуклеотид фосфоэфирной связью с последним нуклеотидом синтезируемой дочерней цепи. При этом отделяется пирофосфат, включающий два остатка фосфорной кислоты, который потом расщепляется на отдельные фосфаты. Реакция отщепления пирофосфата в результате гидролиза энергетически выгодна, так как связь между первым, который уходит в цепь, и вторым фосфатными остатками богата энергией. Эта энергия используется полимеразой.
Полимераза не только удлиняет растущую цепь, но и способна отсоединять ошибочные нуклеотиды, т. е. обладает корректирующей способностью. Если последний нуклеотид, который должен быть присоединен к новой цепи, не комплементарен матричному, то полимераза его удалит.
ДНК-полимераза может присоединять нуклеотид только к -OH группе, находящейся при 3-м атоме углерода дезоксирибозы. Таким образом цепь синтезируется только со стороны своего 3´-конца. То есть синтез новой цепи ДНК идет в направлении от 5´- к 3´-концу. Поскольку в двуцепочечной молекуле ДНК цепи антипараллельны, то процесс синтеза по материнской, или матричной, цепи идет в обратном направлении – от 3´- к 5´-концу.
Поскольку цепи ДНК антипараллельны, а синтез новой цепи возможен только в направлении 5´→3´, то в репликационной вилке дочерние цепи будут синтезироваться в разных направлениях.
На матрице 3´→5´ сборка новой полинуклеотидной последовательности происходит по большей части непрерывно, так как эта цепь синтезируется в направлении 5´→3´. Антипараллельная матрица характеризуется 5´→3´ направлением, поэтому синтез дочерней цепи по ходу движения вилки здесь не возможен. Здесь он был бы 3´→5´, но ДНК-полимера не может присоединять к 5´-концу.
Поэтому синтез на матрице 5´→3´ выполняется небольшими участками — фрагментами Оказаки (названы в честь открывшего их ученого). Каждый фрагмент синтезируется в обратном ходу образования вилки направлении, что обеспечивает соблюдение правила сборки от 5´- к 3´-концу.
Другим «недостатком» полимеразы является то, что она не может сама начать синтез участка дочерней цепи. Причина этого кроется в том, что ей необходим -OH-конец нуклеотида, уже соединенного с цепью. Поэтому необходима затравка, или праймер. Им выступает короткая молекула РНК, синтезируемые ферментом РНК-праймазой и спаренная с матричной цепью ДНК. Синтез каждого участка Оказаки начинается со своей РНК-затравки. Та цепь, которая синтезируется непрерывно, обычно имеет один праймер.
После удаления праймеров и застраивания брешей ДНК-полимеразой отдельные участки дочерней цепи ДНК сшиваются между собой ферментом ДНК-лигазой.
Непрерывная сборка идет быстрее, чем фрагментарная. Поэтому одна из дочерних цепей ДНК называется лидирующей, или ведущей, вторая — запаздывающей, или отстающей.
У прокариот репликация протекает быстрее: примерно 1000 нуклеотидов в секунду. В то время как у эукариот только около 100 нуклеотидов. Количество нуклеотидов в каждом фрагменте Оказаки у эукариот составляет примерно до 200, у прокариот — до 2000.
У прокариот кольцевые молекулы ДНК представляют собой один репликон. У эукариот каждая хромосома может содержать множество репликонов. Поэтому синтез начинается в нескольких точках, одновременно или нет.
Ферменты и другие белки репликации действуют совместно, образуя комплекс и двигаясь по ДНК. Всего в процессе участвует около 20 разных белков, здесь были перечислены лишь основные.
Источник
Репликация
1. Когда происходит репликация? – В синтетической фазе интерфазы, задолго до деления клетки. Период между репликацией и профазой митоза называется постсинтетическая фаза интерфазы, в нем клетка продолжает расти и проверяет, правильно ли произошло удвоение.
2. Если до удвоения было 46 хромосом, то сколько будет после удвоения? – Количество хромосом при удвоении ДНК не изменяется. До удвоения у человека 46 одинарных хромосом (состоящих из одной двойной цепочки ДНК), а после удвоения – 46 двойных хромосом (состоящих из двух одинаковых двойных цепочек ДНК, соединенных между собой в центромере).
3. Зачем нужна репликация? – Чтобы во время митоза каждая дочерняя клетка могла получить свою копию ДНК. При митозе каждая из 46 двойных хромосом делится на две одинарные; получается два набора по 46 одинарных хромосом; эти два набора расходятся в две дочерние клетки.
Три принципа строения ДНК
Полуконсервативность – каждая дочерняя ДНК содержит одну цепочку из материнской ДНК и одну новосинтезированную.
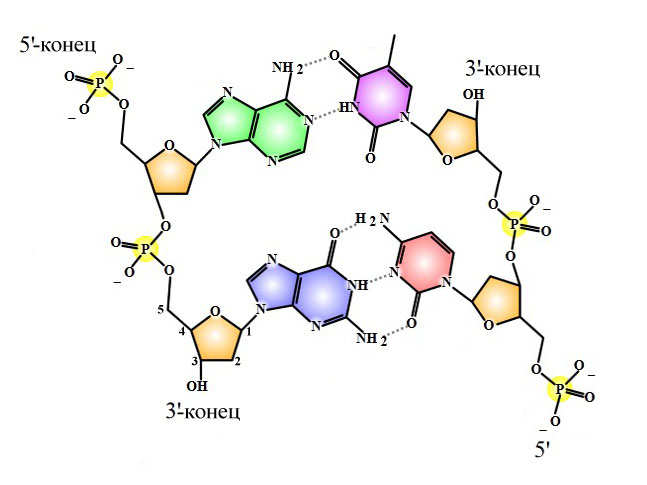
Комплементарность – АТ/ЦГ. Напротив аденина одной цепи ДНК всегда стоит тимин другой цепи ДНК, напротив цитозина всегда стоит гуанин.
Антипараллельность – цепочки ДНК лежат друг к другу противоположными концами. Эти концы не изучают в школе, поэтому чуть подробнее (и далее – в дебри).
Мономером ДНК является нуклеотид, центральной частью нуклеотида – дезоксирибоза. У неё 5 атомов углерода (на ближайшем рисунке у левой нижней дезоксирибозы атомы пронумерованы). Смотрим: к первому атому углерода присоединяется азотистое основание, к пятому – фосфорная кислота данного нуклеотида, третий атом готов присоединить фосфорную кислоту следующего нуклеотида. Таким образом, у любой цепочки ДНК есть два конца:
- 5′-конец, на нем располагается фосфорная кислота;
- 3′-конец, на нем располагается рибоза.
Правило антипараллельности состоит в том, что на одном конце двойной цепи ДНК (например, на верхнем конце ближайшего рисунка) одна цепь имеет 5′-конец, а другая 3′-конец. Для процесса репликации важно, что ДНК-полимераза может удлинять только 3′-конец. Цепочка ДНК может расти только своим 3′-концом.
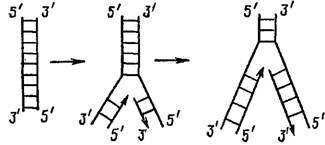
На этом рисунке процесс удвоения ДНК идет снизу вверх. Видно, что левая цепочка растет в том же направлении, а правая – в противоположном.
На следующем рисунке вверхняя новая цепочка («ведущая цепь») удлиняется в том же направлении, в котором происходит удвоение. Нижняя новая цепочка («отстающая цепь») не может удлиняться в том же направлении, потому что там у нее 5′-конец, который, как мы помним, не растёт. Поэтому нижняя цепочка растет с помощью коротких (100-200 нуклеотидов) фрагментов Оказаки, каждый из которых растет в 3′-направлении. Каждый фрагмент Оказаки растет от 3′-конца праймера («РНК-затравки», на рисунке праймеры красные).
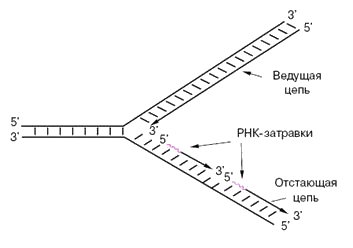
Ферменты репликации
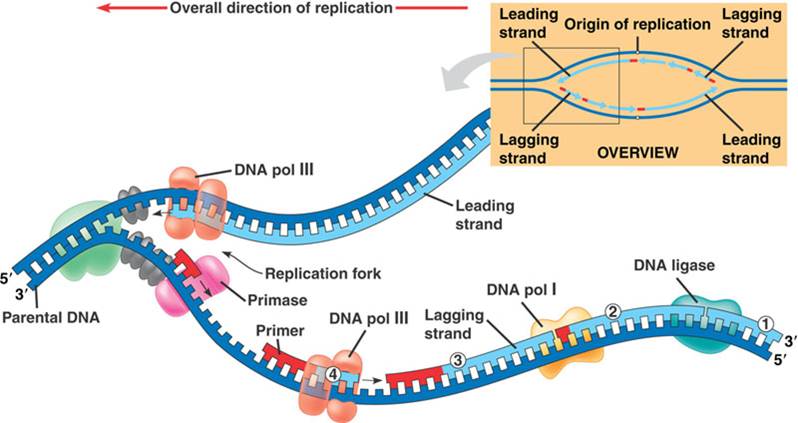
Overall direction of replication – направление, в котором происходит удвоение ДНК.
Parental DNA – старая (материнская) ДНК.
Зеленое облако рядом с надписью «Parental DNA» – фермент хеликаза, который разрывает водородные связи между азотистыми основаниями старой (материнской) цепочки ДНК.
Серые овальчики на только что оторванных друг от друга цепочках ДНК – дестабилизирующие белки, которые не дают цепочкам ДНК соединиться.
DNA pol III – ДНК-полимераза, которая присоединяет новые нуклеотиды к 3′-концу верхней (лидирующей, синтезирующейся неприрывно) цепочки ДНК (Leading strand).
Primase – фермент праймаза, которая делает праймер (красную деталь от Лего). Теперь считаем праймеры слева направо:
- первый праймер еще недоделан, его как раз сейчас делает праймаза;
- от второго по счету праймера ДНК-полимераза строит ДНК – в направлении, противоположном направлению удвоения ДНК, но зато в направлении 3′-конца;
- от третьего по счету праймера цепочка ДНК уже построена (Lagging strand), она подошла вплотную к четвертому по счету праймеру;
- четвертый по счету праймер короче всех, потому что ДНК-полимераза (DNA pol I) удаляет его (он же РНК, в ДНК ему делать нечего, от него нам был нужен только правильный конец) и заменяет на ДНК;
- пятого праймера на рисунке уже нет, он вырезан полностью, на его месте остался разрыв. ДНК-лигаза (DNA ligase) сшивает этот разрыв, чтобы нижняя (отстающая) цепочка ДНК была целой.
На суперкартине не обозначен фермент топоизомераза, но дальше а тестиках он будет фигурировать, так что скажем и про него пару слов. Вот вам веревка, состоящая из трех больших жил. Если три товарища возьмутся за эти три жилы и начнут тянуть их в три разные стороны, то очень скоро веревка перестанет расплетаться и завьется в тугие петли. С ДНК, которая представляет собой двухжильную веревку, могло бы произойти то же самое, если бы не топоизомераза. 
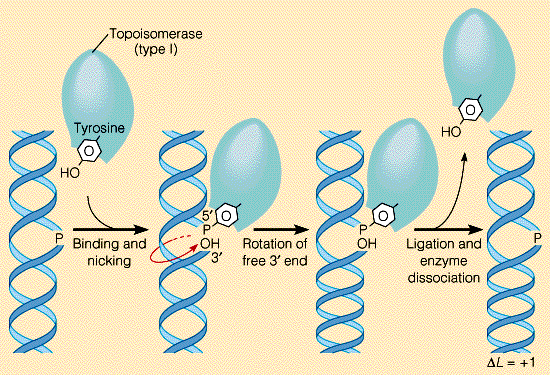
Топоизомереза разрезает одну из двух нитей ДНК, после чего (второй рисунок, красная стрелка) ДНК проворачивается вокруг одной из своих цепей, так что тугие петли не образуются (топологический стресс снижается).
Концевая недорепликация
Из суперкартины с ферментами репликации понятно, что на месте, оставшемся после удаления праймера, ДНК-полимераза достраивает следующий по счету фрагмент Оказаки. (Правда понятно? Если что, фрагменты Оказаки на суперкартине обозначены цифрами в кружочках.) Когда репликация на суперкартине дойдет до своего логического (левого) конца, то у последнего (крайнего левого) фрагмента Оказаки не будет «следующего», поэтому некому будет достроить ДНК на пустом месте, получившемся после удаления праймера.
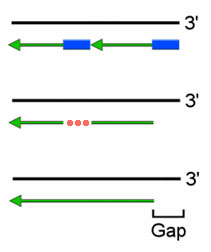
Вот вам еще рисунок. Черная цепочка ДНК – старая, материнская. Удвоение ДНК, в отличие от суперкартины, происходит слева направо. Поскольку у новой (зеленой) ДНК справа 5′-конец, то она является отстающей и удлиняется отдельными фрагметами (Оказаки). Каждый фрагмент Оказаки растет от 3′-конца своего праймера (синего прямоугольника). Праймеры, как мы помним, удаляются ДНК-полимеразой, которая на этом месте достраивает следующий фрагмент Оказаки (этот процесс обозначен красным многоточием). На конце хромосомы некому заделать этот участок, так как нету следующего фрагмента Оказаки, там уже пустое место (Gap). Таким образом, после каждой репликации у дочерних хромосом укорачиваются оба 5′-конца (концевая недорепликация).
Чтобы концевая недорепликация не приводила к печальным последствиям, на концах хромосом имеются участки, не несущие наследственной информации – теломеры. Их укорочение не приносит вреда; у человека они рассчитаны примерно на 60 репликаций. Больше 60 раз (число Хейфлика) клетки человека поделиться не могут, поскольку концевая недорепликация начинает затрагивать гены.
Стволовые клетки (в коже, красном костном мозге, семенниках) должны делиться гораздо больше, чем 60 раз. Поэтому в них функционирует фермент теломераза, который после каждой репликации удлиняет теломеры. Теломераза удлиняет выступающий 3′-конец ДНК, так что он увеличивается до размера фрагмента Оказаки. После этого праймаза синтезирует на нем праймер, и ДНК-полимераза удлиняет недореплицированный 5′-конец ДНК.
Тестики
1. Репликация — это процесс, в котором:
А) происходит синтез транспортных РНК;
Б) происходит синтез (копирование) ДНК;
В) рибосомы узнают антикодоны;
Г) образуются пептидные связи.
2. Соотнесите функции ферментов, участвующих в репликации прокариот, с их названиями.
| Ферменты | Функции |
| 1) ДНК геликаза | а) синтез РНК-праймера на отрезке прерывной репликации |
| 2) Праймаза | б) раскрутка двуцепочечной ДНК |
| 3) ДНК полимераза I 3´→5´-нуклеазная активность | в) удаление праймера РНК |
| 4) ДНК полимераза I 5´→3´-нуклеазная активность | г) сшивание пробелов между фрагментами Оказаки |
| 5) ДНК лигаза | д) удаление неспаренных нуклеотидов |
| 6) Топоизомераза | е) снижение топологического стресса при расщеплении двуцепочечной ДНК |
3. Во время репликации в эукариотических клетках удаление праймеров
А) осуществляется ферментом только с ДНК-азной активностью
Б) образует фрагменты Оказаки
В) происходит только в отстающих цепях
Г) происходит только в ядре
4. Если Вы проэкстрагируете ДНК бактериофага fX174, вы обнаружите, что в его составе находится 25% A, 33% T, 24% G, и 18% C. Как Вы могли бы обьяснить эти результаты?
А) Результаты эксперимента неправильные; где-то произошла ошибка.
Б) Можно было бы допустить, что процентное содержание A приблизительно равно таковому T, что также справедливо для C и G. Следовательно, правило Чаргаффа не нарушается, ДНК является двуцепочечной и реплицируется полуконсервативно.
В) Поскольку процентные соотношения A и T и, соответственно, C и G различные, ДНК представляет собой одну цепь; она реплицируется при помощи особенного фермента, следующего особенному механизму репликации с одной цепью в качестве матрицы.
Г) Поскольку ни A не равно T, и ни G не равно C, то ДНК должна быть одноцепочечной, она реплицируется путем синтеза комплементарной цепи и использованием этой двуцепочечной формы как матрицы.
5. Диаграмма относится к репликации двуцепочечной ДНК. Для каждого из квадратов I, II, III выберите один фермент, который функционирует на этом участке.
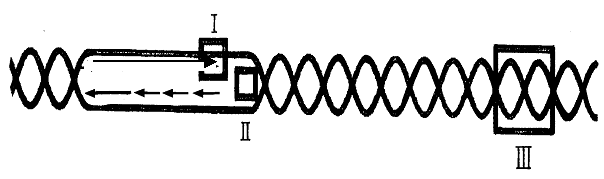
А) Теломераза
Б) ДНК-топоизомераза
В) ДНК-полимераза
Г) ДНК-геликаза
Д) ДНК-лигаза
6. Культура бактерий из среды с легким изотопом азота (N-14) перенесли в среду, содержащую тяжелый изотоп (N-15) на время, соответствующее одному делению, а затем вернули в среду с легким изотопом азота. Анализ состава ДНК бактерий после периода, соответствующего двум репликациям, показал:
| Варианты ответа | ДНК | ||
| легкая | средняя | тяжелая | |
| А | 3/4 | 1/4 | — |
| Б | 1/4 | 3/4 | — |
| В | — | 1/2 | 1/2 |
| Г | 1/2 | 1/2 | — |
7. Одно редкое генетическим заболевание характеризуется иммунодефицитом, отставанием в умственном и физическом развитии и микроцефалией. Предположим, что в экстракте ДНК пациента с этим синдромом вы обнаружили почти одинаковые количества длинных и очень коротких отрезков ДНК. Какой фермент у этого пациента наиболее вероятно отсутствует/дефектный?
А) ДНК-лигаза
Б) Топоизомераза
В) ДНК-полимераза
Г) Геликаза
8. Молекула ДНК, представляет собой двойную спираль, содержащую четыре различных типа азотистых оснований. Какое из следующих утверждений в отношении как репликации, так и химического строения ДНК, является правильным?
A) Последовательности оснований двух цепей одни и те же.
B) В двойной цепи ДНК содержание пуринов равно содержанию пиримидинов.
C) Обе цепи синтезируются в направлении 5’→3’ непрерывно.
D) Присоединение первого основания вновь синтезируемой нуклеиновой кислоты катализируется ДНК-полимеразой.
E) Активность ДНК-полимеразы по исправлению ошибок осуществляется в направлении 5’→3’.
9. Большинство ДНК-полимераз обладает также активностью:
А) лигазной;
Б) эндонуклеазной;
В) 5′-экзонуклеазной;
Г) 3′-экзонуклеазной.
10. ДНК-хеликаза — это ключевой фермент репликации ДНК, раскручивающий двуцепочечную ДНК до одноцепочечной. Ниже описан эксперимент, посвященный выяснению свойств этого фермента.
Линейная одноцепочечная ДНК (ssDNA) длиной 6000 нуклеотидов была гибридизована с короткой (300 нуклеотидов) комплементарной одноцеаочечной ДНК, меченой радиоактивными нуклеотидами (a). Эта гибридизованная ДНК была обработана одним из трех способов: ДНК-хеликазой, кипячением без геликазы или прокипяченной геликазой. Затем образцы ДНК были подвергнуты электрофорезу в агарозном геле. На рисунке b показаны полосы ДНК, которые можно было обнаружить в геле при помощи авторадиографии. (Предположено, что необходимая для этой энзиматической реакции энергия АТФ была предоставлена во время обработки ДНК-хеликазой). 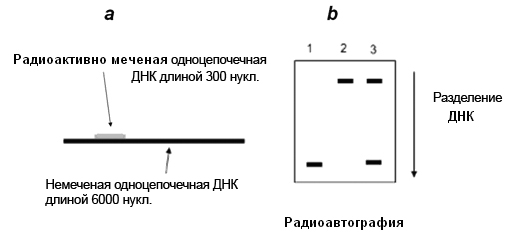
Какое из следующих утверждений относительно этого эксперимента является правильным?
А) Полоса, появляющаяся в верхней части геля, является только ssДНК, величиной 6,3 kb.
Б) Полоса, появляющаяся в нижней части геля, это меченная 300bp ДНК.
В) Если гибридизованную ДНК обработать только ДНК хеликазой и довести реакцию до конца, расположение полос выглядит так, как изображено на дорожке 3 на рисунке b.
Г) Если гибридизованную ДНК обработать только кипячением без обработки хеликазой, расположение полос выглядит как изображено на дорожке 2 на рисунке b.
Д) Если гибридизованную ДНК обработать только прокипяченной хеликазой, расположение полос выглядит как изображено на дорожке 1 на рисунке b.
[1] — окружная олимпиада 2001
[2] — всероссийская олимпиада 2001
[3, 4, 5, 8] — международная олимпиада 2001
[6] — международная олимпиада 1991
[7] — международная олимпиада 2008
[9] — окружная олимпиада 2008
[10] — международная олимпиада 2010
Полные тексты этих олимпиад можно найти здесь.
Источник



