
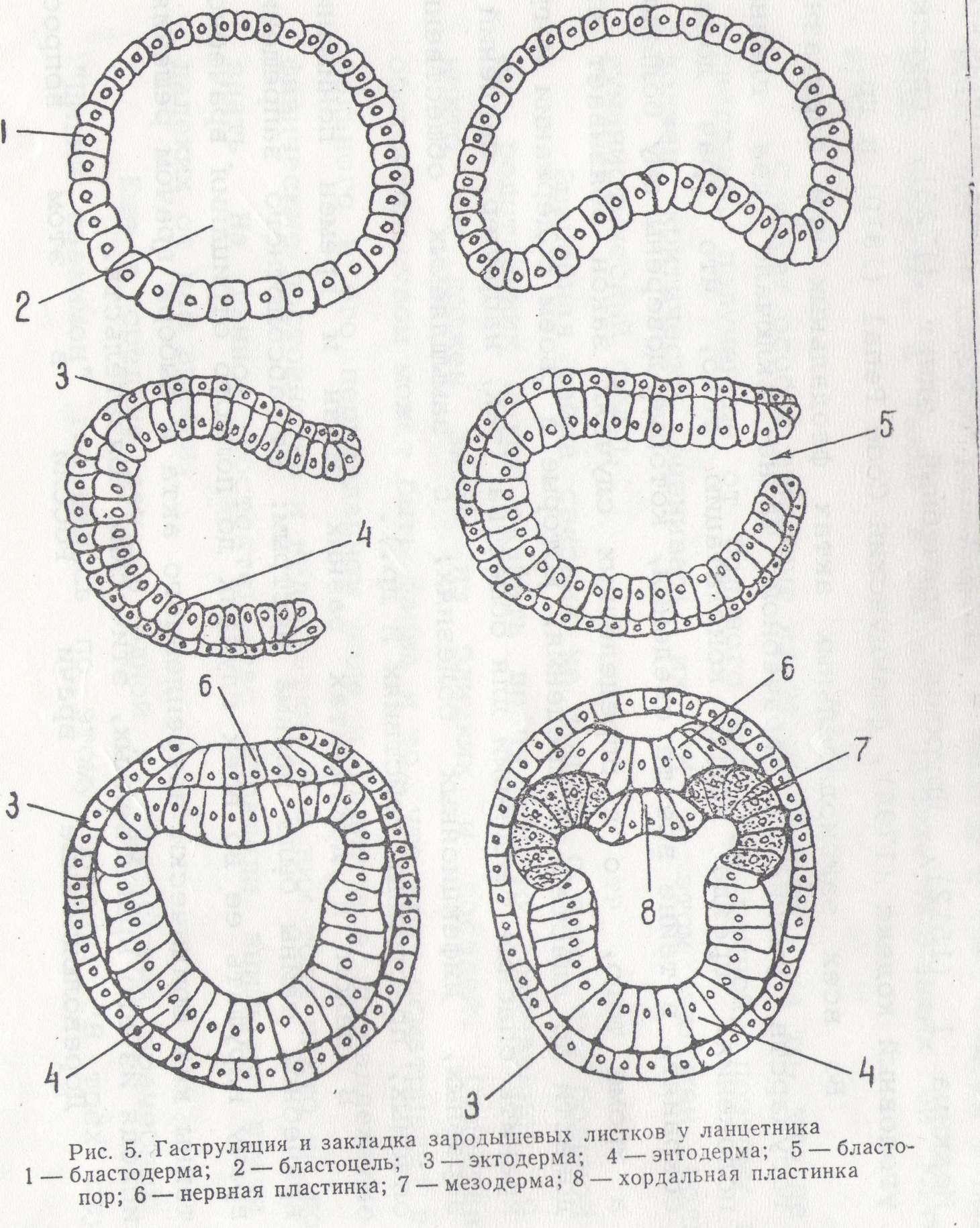
Гаструляция у ланцетника
Гаструляция у ланцетника представляет удобную модель для изучения общих закономерностей этого процесса у типа хордовых. Этот процесс, как было показано ранее, у ланцетника, происходит по типу инвагинации. На ранней стадии гаструлы обнаруживается два зародышевых листка: эктодермальный, образованный из крыши бластулы, и энтодермальный— из дна бластулы. Гастроцель сообщается с внешней средой при помощи бластопора.
Наступает следующий этап развития зародыша — образование третьего зародышевого листка—мезодермы. Клетки дорзальной губы бластопора, быстро размножаясь, погружаются в составе энтодермы и на дорзальной ее стороне образуют хордальную пластинку— зачаток будущей хорды. Мелкие клетки вентральной и боковых губ бластопора также врастают в энтодермальный листок и располагаются в его составе по сторонам от хордальной пластинки.
Двуслойный зародыш приобретает вытянутую форму, имея в своем составе материал трех зародышевых листков. Материал мезодермы занимает дорзальную часть внутреннего листка. Из первичной эктодермы, расположенной над хордальной пластинкой, выделяется нервная пластинка. Последняя погружается и по длине зародыша образует нервный желобок. Нервный желобок, замыкаясь кверху, образует нервную трубку.
Хордальная пластинка выходит из состава первичного внутреннего листка, замыкается книзу и образует плотный клеточный тяж — хорду.
Одновременно мелкоклеточный материал, находившийся в первичном внутреннем листке по сторонам от хорды, выпячивается в дорзальном направлении в виде двух карманов. Вначале эти карманы открываются в гастроцель, а затем отделяются в виде двух замкнутых складок, расположенных вдоль гаструлы.
Так образуется третий зародышевый листок — мезодерма. После выделения хорды и мезодермы материал первичного внутренного листка смыкается, образуя энтодерму — внутренний зародышевый листок (рис. 5).
ГАСТРУЛЯЦИЯ у птиц
Гаструляция у птиц представляет довольно близкую модель процессам, имеющим место у высших хордовых, в том числе и у человека.
У птиц в результате дискоидального (меробластического) дробления образуется бластодиск. Он состоит из клеток неправильной формы, плотно прилегающих друг к другу и расположенных на огромной массе нераздробившегося желтка. Под влиянием ферментов клеток бластодиска часть желтка разжижается и образуется полость, заполненная жидкостью — бластоцель. Крыша этой бластулы представлена бластодиском, дно — нераздробившейся массой желтка. Бластоцель — подзародышевая полость.
Затем наступает гаструляция, состоящая из двух фаз. Первая фаза начинается до откладывания яйца и заключается в обособлении энтодермы путем деляминации клеток бластодиска. В результате возникает двуслойный зародыш. Верхний слой клеток дискобластулы приобретает призматическую форму и располагается в один правильный ряд. Нижний слой клеток дискобластулы сохраняет округлую или неправильную форму и лежит беспорядочно на желтке. Между клетками верхнего и нижнего слоя возникает полость — гастроцель.
Вторая фаза гаструляции начинается во второй половине первых суток инкубации. Бластодиск разрастается, в центре его выделяется зародышевый щиток, из которого в дальнейшем развивается тело зародыша. Окружающая зародышевый щиток часть дискобластулы представляет внезародышевый материал, в котором различают светлое и темное поле. Светлое поле расположено вокруг зародышевого щитка и состоит из клеток, отделившихся от желтка подзародышевой полостью в силу частичного использования желтка зародышем. Темное поле занимает периферию дискобластулы и состоит из клеток, плотно прилегающих к желтку и растущих по его поверхности. К этому моменту резко выражено разделение клеточного материала на зародышевую (зародышевый щиток) и внезародышевую (светлое и темное поле) часть.
В течение 24 часов насиживания или инкубации на зародышевом щитке в результате размножения клеток наблюдается перемещение их спереди назад по периферии зародышевого щитка. Оба потока клеток встречаются в центре у заднего конца зародышевого щитка, сливаются и перемещаются кпереди посредине зародышевого щитка. В результате образуется утолщенный тяж клеток, получивший название первичной полоски.
На переднем конце первичной полоски формируется утолщение— первичный узелок. В дальнейшем часть бластомеров, расположенных кпереди от первичного узелка, перемещается к последнему, подворачивается под эктодерму и формирует растущий кпереди между эктодермой и энтодермой головной или хордальный вырост — зачаток хорды.
Бластомеры задней половины зародышевого щитка смещаются к первичной полоске и через эту область погружаются под эктодерму, располагаясь в полости между эктодермой и энтодермой. Этот материал образует зачаток мезодермы — третьего зародышевого листка. Клетки хордального выроста размножаются и. перемещаются, занимая центральное положение между эктодермой
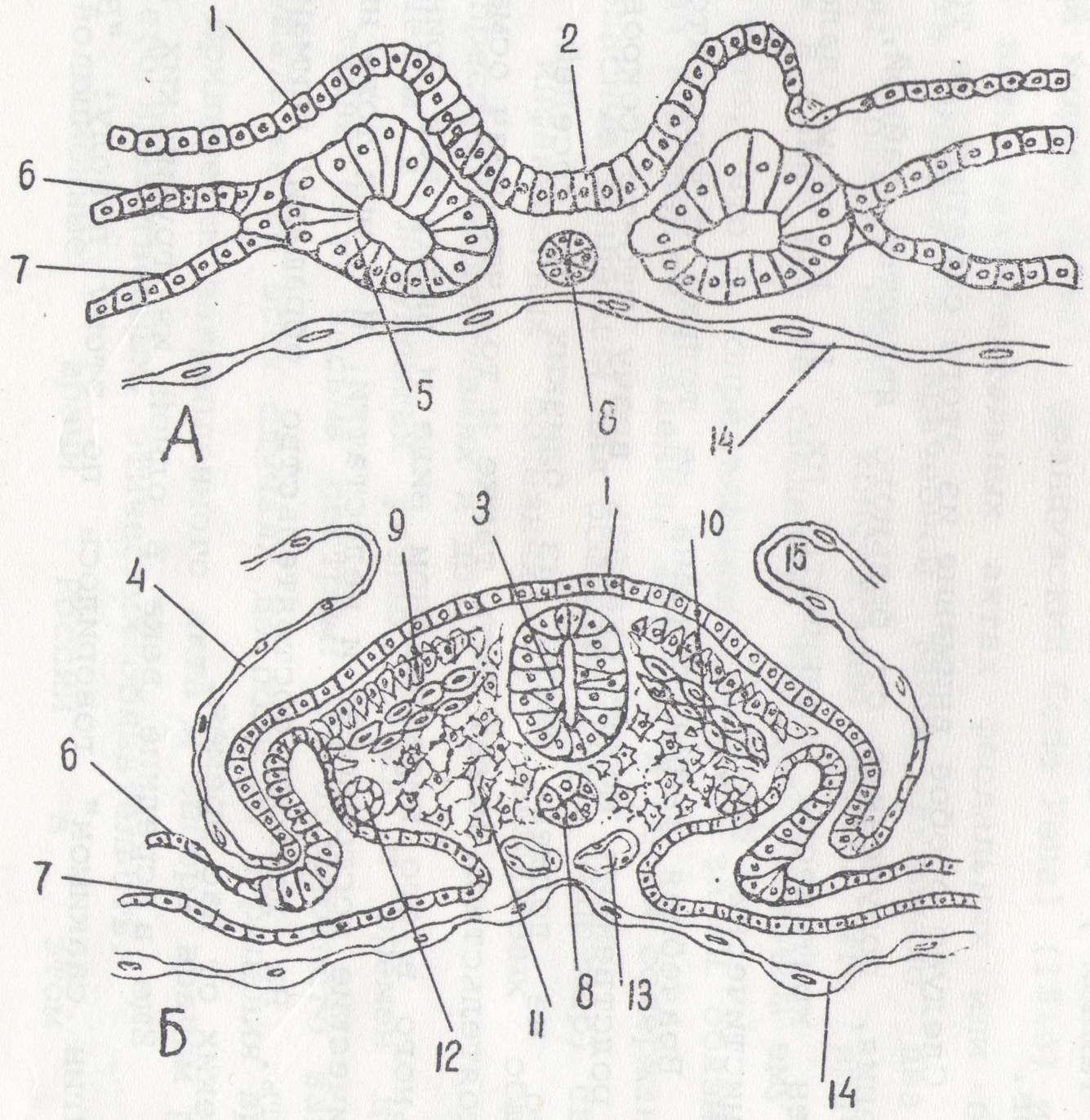
Рис.%6. Развитие комплекса осевых органов у птиц 1—эктодерма; 2 —нервный желобок; 3 — нервная трубка; 4 — амнион; 5 — сомиты; 6 — париетальный листок сплахнотома; 7 — висцеральный листок спланхнотома; 8 — хорда; 9 — дерматом; 10 — миотом; 11 — склеротом; 12 — нефротом; 13 — аорта; 14 — энтодерма; 15 — амниотическая складка и энтодермой в передней половине зародышевого щитка Вслед за этим клетки мезодермы также перемещаются в переднюю половину зародышевого щитка, располагаясь между эктодермой и энтодермой по бокам от хордального выроста. Это парахордальная мезодерма. Ее можно подразделить на дорзальную и вентральную.
По мере того как клетки первичной полоски уходят в закладки хорды и мезодермы на поверхности остается материал эктодермы и входящей в нее нервной пластинки. Таким образом, первичная полоска и головной узелок по своему значению соответствуют бластопору ланцетника, т. к. в этом месте проходит перемещение зародышевого материала, приводящее к образованию 3-зародышевых листков. Наиболее глубоко расположен зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы.
К концу гаструляции в теле зародыша имеются все зачатки, из которых возникает типичный для хордовых комплекс осевых органов (рис. 6.).
Источник
Каким способом происходит гаструляция у ланцетника
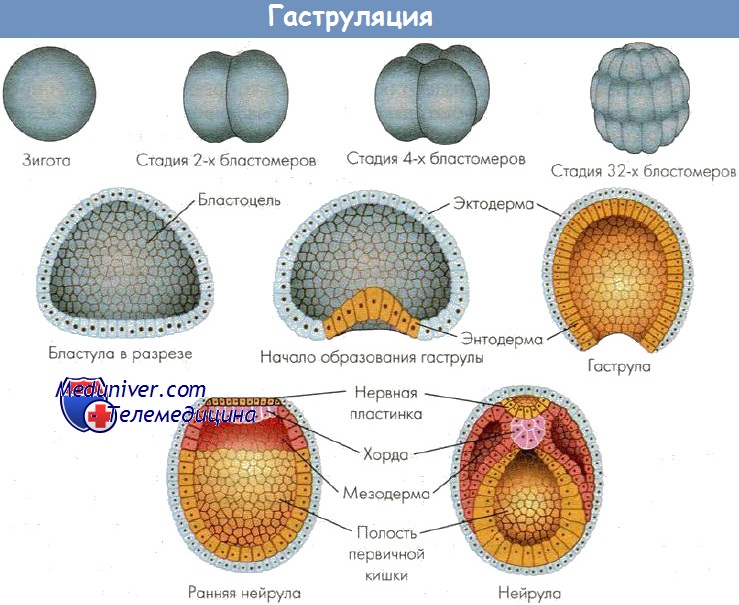
Гаструляция — результат активного деления клеток, роста и направленных перемещений (миграций) клеточных потоков с формированием многослойного зародыша, или гаструлы, (возникновением послойно расположенных, отделенных друг от друга отчетливой щелью, зародышевых листков: наружного — эктодермы, среднего — мезодермы, внутреннего — энтодермы). Перемещение клеток происходит в строго определенной области зародыша — в области серого серпа. Последний был описан В. Ру в 1888 г. В оплодотворенном яйце амфибии серый серп выявляется как окрашенная область на стороне, противоположной проникновению спермия. В этом месте, как полагают, локализуются факторы, необходимые для гаструляции.
У разных представителей позвоночных гаструляция совершается несколькими основными способами: путем инвагинации (впячивания), иммиграции (перемещения части клеток внутрь зародыша), эпиболии (обрастания), деламинации (расщепления). Способы гаструляции зависят от типа яйцеклетки. При любом способе гаструляции в качестве ведущих сил выступают неравномерная пролиферация клеток в разных частях зародыша, уровень обменных процессов в клетках, расположенных в разных частях зародыша, активность амебоидных движений клеток, а также индуктивные факторы (белки, нуклеопротеиды, стероиды и др.). В результате гаструляции обособляются основные зачатки органов и тканей.
Следующим периодом эмбриогенеза является гисто- и органогенез — дифференцировка различных тканей и органов организма из материала зародышевых листков и эмбриональных зачатков.
В результате гаструляции возникает многослойный зародыш. Несмотря на различные способы гаструляции после выделения материала зародышевых листков по оси зародыша находится материал хорды, который подстилает нервную пластинку, слева и справа от хорды располагается материал мезодермы. Все это характеризует осевой комплекс зачатков. В дальнейшем происходит формирование зачатков органов, представляющих собой пространственно локализованные группы стволовых клеток — источников развития тканей. Закономерности дифференцировки клеточного материала зачатков можно проследить в эмбриогенезе наиболее изученных животных.
Ланцетник. Развитие ланцетника.
Классическим объектом эмбриологических исследований ланцентник, подробно изученный А.О. Ковалевским. Ланцетник является представителем класса хордовых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне в теплых морях. Свое название получил из-за формы, напоминающей ланцет (хирургический инструмент с обоюдоострым лезвием, современный скальпель).
Яйцеклетка ланцетника олиго- и изолецитальная, размером 110 мкм, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Дробление зиготы полное, почти равномерное, синхронное и заканчивается образованием бластулы. В результате чередования меридианальных и широтных борозд дробления формируется однослойная бластула с полостью, заполненной жидкостью — бластоцелем. Бластула сохраняет полярность, ее дно представляет собой вегетативную, а крыша — анимальную части; между ними находится краевая зона.
При гаструляции происходит впячивание вегетативной части бластулы в анимальную. Впячивание постепенно углубляется и, наконец, формируется двустенная чаша с широко зияющим отверстием, ведущим в новообразованную полость зародыша. Такой способ гаструляции называется инвагинацией. Так бластула превращается в гаструлу. В ней материал зародыша оказывается дифференцированным на наружный листок — эктодерму, и внутренний — энтодерму. Полость чаши именуется гастроцелем, или полостью первичной кишки, которая сообщается с внешней средой посредством бластопора, что соответствует заднепроходному отверстию. В бластопоре различают дорсальную, вентральную и две латеральных губы. В результате инвагинации центр тяжести зародыша смещается, и зародыш поворачивается бластопором кверху. Постепенно происходит смыкание краев бластопора и удлинение зародыша. Топография клеток в составе губ бластопора определяет развитие разных частей зародыша. При гаструляции из внутреннего листка гаструлы обособляются хорда и мезодерма, которые располагаются между экто- и энтодермой. Завершается гаструляция образованием осевого комплекса зачатков и дальше — обособлением зачатков органов. Хорда индуцирует развитие нервной трубки из материала дорсальной эктодермы. Эта часть эктодермы утолщается, образуется нервная пластинка (нейроэктодерма), которая прогибается по средней линии и превращается в желобок.
Края желобка постепенно смыкаются в нервную трубку. Оставшаяся часть эктодермы — кожная, срастается над нервной трубкой. Однако на самом переднем и заднем концах зародыша нервная трубка некоторое время сообщается с внешней средой с помощью двух отверстий — нейропоров. В последующем происходит разделение мезодермы на спинные сегменты — сомиты, количество которых нарастает от 15 пар до 60-65 пар у взрослого ланцетника. Часть латерально расположенной мезодермы не сегментируется и расщепляется на наружный (париетальный) и внутренний (висцеральный) листки спланхнотома. Эти листки растут между экто- и энтодермой и, достигнув середины на вентральной стороне зародыша под кишечной трубкой, срастаются, формируя единую вторичную полость — целом. На переднем конце зародыша возникает углубление (ротовая бухта), растущее по направлению к переднему отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепого конца кишечной трубки происходит апоптоз клеток и возникает сообщение кишки с внешней средой. Аналогичный процесс протекает на заднем конце зародыша. По бокам головного отдела зародыша также возникает контакт кожной эктодермы и кишечной энтодермы. В месте этого контакта происходит прорыв. Так полость передней кишки сообщается с внешней средой (формируется жаберный аппарат). После этого зародыш выходит из яйцевой оболочки во внешнюю среду в виде личинки.
Методы маркирования для изучения процессов миграции бластомеров позволили выделить определенные области зародыша на ранних стадиях развития (зиготы — бластулы), которые позднее развиваются в зародышевые листки и эмбриональные зачатки органов и тканей. Эти области были названы презумптивными (предполагаемыми) участками, или зачатками.
Таким образом, на примере развития ланцетника можно наблюдать закономерное перемещение клеток и формирование зародышевых листков, осевого комплекса зачатков и обособление зачатков органов и тканей.
Источник
Гаструляция у ланцетника
Гаструляция у ланцетника представляет удобную модель для изучения общих закономерностей этого процесса у типа хордовых. Этот процесс, как было показано ранее, у ланцетника, происходит по типу инвагинации. На ранней стадии гаструлы обнаруживается два зародышевых листка: эктодермальный, образованный из крыши бластулы, и энтодермальный— из дна бластулы. Гастроцель сообщается с внешней средой при помощи бластопора.
Наступает следующий этап развития зародыша — образование третьего зародышевого листка—мезодермы. Клетки дорзальной губы бластопора, быстро размножаясь, погружаются в составе энтодермы и на дорзальной ее стороне образуют хордальную пластинку— зачаток будущей хорды. Мелкие клетки вентральной и боковых губ бластопора также врастают в энтодермальный листок и располагаются в его составе по сторонам от хордальной пластинки.
Двуслойный зародыш приобретает вытянутую форму, имея в своем составе материал трех зародышевых листков. Материал мезодермы занимает дорзальную часть внутреннего листка. Из первичной эктодермы, расположенной над хордальной пластинкой, выделяется нервная пластинка. Последняя погружается и по длине зародыша образует нервный желобок. Нервный желобок, замыкаясь кверху, образует нервную трубку.
Хордальная пластинка выходит из состава первичного внутреннего листка, замыкается книзу и образует плотный клеточный тяж — хорду.
Одновременно мелкоклеточный материал, находившийся в первичном внутреннем листке по сторонам от хорды, выпячивается в дорзальном направлении в виде двух карманов. Вначале эти карманы открываются в гастроцель, а затем отделяются в виде двух замкнутых складок, расположенных вдоль гаструлы.
Так образуется третий зародышевый листок — мезодерма. После выделения хорды и мезодермы материал первичного внутренного листка смыкается, образуя энтодерму — внутренний зародышевый листок (рис. 5).
ГАСТРУЛЯЦИЯ у птиц
Гаструляция у птиц представляет довольно близкую модель процессам, имеющим место у высших хордовых, в том числе и у человека.
У птиц в результате дискоидального (меробластического) дробления образуется бластодиск. Он состоит из клеток неправильной формы, плотно прилегающих друг к другу и расположенных на огромной массе нераздробившегося желтка. Под влиянием ферментов клеток бластодиска часть желтка разжижается и образуется полость, заполненная жидкостью — бластоцель. Крыша этой бластулы представлена бластодиском, дно — нераздробившейся массой желтка. Бластоцель — подзародышевая полость.
Затем наступает гаструляция, состоящая из двух фаз. Первая фаза начинается до откладывания яйца и заключается в обособлении энтодермы путем деляминации клеток бластодиска. В результате возникает двуслойный зародыш. Верхний слой клеток дискобластулы приобретает призматическую форму и располагается в один правильный ряд. Нижний слой клеток дискобластулы сохраняет округлую или неправильную форму и лежит беспорядочно на желтке. Между клетками верхнего и нижнего слоя возникает полость — гастроцель.
Вторая фаза гаструляции начинается во второй половине первых суток инкубации. Бластодиск разрастается, в центре его выделяется зародышевый щиток, из которого в дальнейшем развивается тело зародыша. Окружающая зародышевый щиток часть дискобластулы представляет внезародышевый материал, в котором различают светлое и темное поле. Светлое поле расположено вокруг зародышевого щитка и состоит из клеток, отделившихся от желтка подзародышевой полостью в силу частичного использования желтка зародышем. Темное поле занимает периферию дискобластулы и состоит из клеток, плотно прилегающих к желтку и растущих по его поверхности. К этому моменту резко выражено разделение клеточного материала на зародышевую (зародышевый щиток) и внезародышевую (светлое и темное поле) часть.
В течение 24 часов насиживания или инкубации на зародышевом щитке в результате размножения клеток наблюдается перемещение их спереди назад по периферии зародышевого щитка. Оба потока клеток встречаются в центре у заднего конца зародышевого щитка, сливаются и перемещаются кпереди посредине зародышевого щитка. В результате образуется утолщенный тяж клеток, получивший название первичной полоски.
На переднем конце первичной полоски формируется утолщение— первичный узелок. В дальнейшем часть бластомеров, расположенных кпереди от первичного узелка, перемещается к последнему, подворачивается под эктодерму и формирует растущий кпереди между эктодермой и энтодермой головной или хордальный вырост — зачаток хорды.
Бластомеры задней половины зародышевого щитка смещаются к первичной полоске и через эту область погружаются под эктодерму, располагаясь в полости между эктодермой и энтодермой. Этот материал образует зачаток мезодермы — третьего зародышевого листка. Клетки хордального выроста размножаются и. перемещаются, занимая центральное положение между эктодермой
Рис.%6. Развитие комплекса осевых органов у птиц 1—эктодерма; 2 —нервный желобок; 3 — нервная трубка; 4 — амнион; 5 — сомиты; 6 — париетальный листок сплахнотома; 7 — висцеральный листок спланхнотома; 8 — хорда; 9 — дерматом; 10 — миотом; 11 — склеротом; 12 — нефротом; 13 — аорта; 14 — энтодерма; 15 — амниотическая складка и энтодермой в передней половине зародышевого щитка Вслед за этим клетки мезодермы также перемещаются в переднюю половину зародышевого щитка, располагаясь между эктодермой и энтодермой по бокам от хордального выроста. Это парахордальная мезодерма. Ее можно подразделить на дорзальную и вентральную.
По мере того как клетки первичной полоски уходят в закладки хорды и мезодермы на поверхности остается материал эктодермы и входящей в нее нервной пластинки. Таким образом, первичная полоска и головной узелок по своему значению соответствуют бластопору ланцетника, т. к. в этом месте проходит перемещение зародышевого материала, приводящее к образованию 3-зародышевых листков. Наиболее глубоко расположен зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы.
К концу гаструляции в теле зародыша имеются все зачатки, из которых возникает типичный для хордовых комплекс осевых органов (рис. 6.).
Источник



