
Какие существуют способы передачи наследственной информации
§ 15. ПЕРЕНОС ГЕНЕТИЧЕСКОЙ ИНФОРМАЦИИ В КЛЕТКЕ:
РЕПЛИКАЦИЯ, ТРАНСКРИПЦИЯ
В процессе размножения клеток генетическая информация передается от одного поколения клеток другому. При этом все клетки получают одинаковую информацию. Это, возможно, вследствие того, что перед делением клетки осуществляется репликация (удвоение) ДНК, в результате образуются две идентичные молекулы ДНК, которые и передаются потомкам. В структуре ДНК заложена способность этой молекулы к копированию.
Закодированная в ДНК генетическая информация реализуется в результате экспрессии генов. Экспрессия генов включает транскрипцию (копирование информации с ДНК на синтезируемую РНК) и последующую трансляцию (синтез на матрице РНК соответствующего белка).
Возможен поток информации и в направлении от РНК к ДНК, этот процесс носит название обратная транскрипция. В то же время информация не передается от белков нуклеиновым кислотам. Однако следует отметить, что белки играют важную роль в осуществлении процессов передачи информации, как между нуклеиновыми кислотами, так и от нуклеиновых кислот к белкам.
Описанные информационные взаимоотношения между ДНК, РНК и белками могут быть представлены в виде схемы:
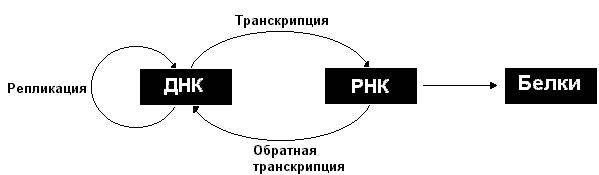
Рассмотрим более подробно процессы репликации, транскрипции и трансляции.
Интересно знать! В 1982 году Р.Д.Пальмитер с коллегами опубликовали фотографию, на которой рядом сидели две мыши. Одна из них была трансгенной, в ее ДНК встроили ген гормона роста крысы, другая была обычной мышкой. Трансгенная мышь в сравнении с обычной имела в два раза большие размеры. Причиной тому была экспрессия гена гормона роста, в результате которой у мыши синтезировался соответствующий гормон, который и определил ее гигантские размеры.
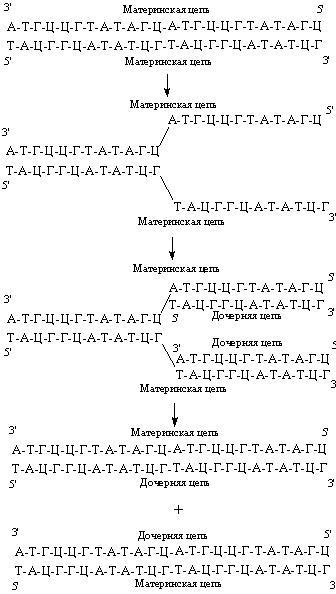
Рис. 44. В результате репликации образуются две дочерние молекулы ДНК.
Репликация ДНК
В основе репликации, или удвоения, ДНК лежит принцип комплементарности. Репликация начинается с разделения цепей, каждая из которых становится матрицей, определяющей нуклеотидную последовательность новой комплементарной цепи (рис. 44). В результате репликации образуются две дочерние молекулы ДНК, нуклеотидные последовательности которых идентичны между собой и с материнской молекулой ДНК. При этом две дочерние молекулы ДНК содержат одну вновь синтезированную (дочернюю) и одну материнскую цепи ДНК.
Субстратом для синтеза ДНК являются дезоксинуклеотид-5’-трифосфаты: дАТФ, дГТФ, дЦТФ и дТТФ. Уравнение этой реакции в простейшей виде выглядит так:
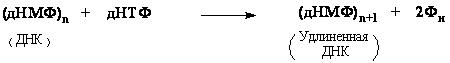
Данную реакцию катализирует фермент ДНК-полимераза. Она последовательно добавляет нуклеотиды к 3’-концу полинуклеотидной цепи. Следовательно, синтез новой цепи происходит в направлении от 5’-конца к 3’-концу. ДНК-полимераза может только наращивать цепь ДНК, начать же синтез ДНК с нуля она не может, т.е. для ее работы требуется цепь-затравка. ДНК-полимераза способна удлинять цепь только в присутствии цепи, играющей роль матрицы (рис. 45.). Нуклеотиды присоединяются к цепи-затравке в соответствии с принципом комплементарности, напротив аденина всегда будет встроен тимин, а напротив гуанина – цитозин.
Итогом репликации является образование двух дочерних молекул ДНК, являющихся точными копиями материнской. Благодаря этому возможна передача идентичной генетической информации от клетки к клетке.
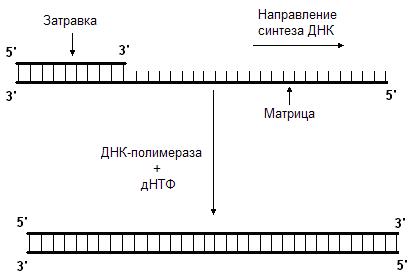
Рис. 45. ДНК-полимераза наращивает цепь в направлении 5’ 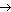 3’.
3’.
Для этого ей нужны матрица и затравка
Транскрипция
В результате транскрипции происходит синтез цепи РНК, нуклеотидная последовательность которой комплементарна одной из цепей ДНК; при этом генетическая информация, содержащаяся в ДНК, передается молекуле РНК. В результате транскрипции образуются три класса РНК: иРНК, тРНК, рРНК. Поскольку в основе транскрипции лежит принцип комплементарности, этот процесс имеет определенное сходство с репликацией. Однако между ними существует и важное различие. Если в процессе репликации копируется вся молекула ДНК, то при транскрипции транскрибируется только ее незначительная часть. Катализирует синтез РНК фермент РНК-полимераза. Транскрибируемый участок ДНК ограничен со стороны 3’-конца промотором, – участком с которым связывается РНК-полимераза, со стороны 5’-конца – терминатором – участком, в котором прерывается синтез РНК. Последовательность ДНК, ограниченная промотором и терминатором, представляет собой единицу транскрипции – транскриптон (рис. 46).
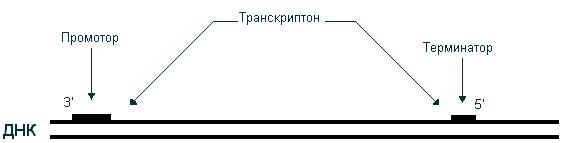
Рис. 46. Транскриптон
РНК-полимераза в качестве субстратов для синтеза РНК использует рибонуклеозид-5’-фосфаты (АТФ, ГТФ, ЦТФ и УТФ). Она удлиняет цепь, присоединяя нуклеотиды к 3’-концу, т.е. так же, как и ДНК-полимераза, наращивает цепь в направлении5’®3’. Этот фермент требует для своей работы в качестве матрицы одну из цепей ДНК. Из двух цепей ДНК транскрибируется только одна. В отличие от ДНК-полимеразы, РНК-полимераза не нуждается в затравке. Удлинение цепи РНК описывается уравнением:

Нуклеотиды присоединяются к цепи в соответствии с принципом комплементарности. Напротив аденина матричной цепи ДНК в растущую цепь РНК всегда будет встроен урацил, напротив гуанина – цитозин, напротив тимина – аденин и напротив цитозина – гуанин соответственно. Т.о., образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК (рис. 47).
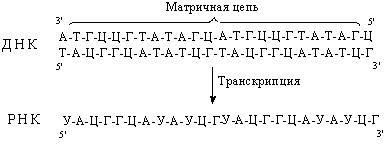
Рис. 47. Образовавшаяся в результате транскрипции РНК комплементарна матричной цепи ДНК
В процессе транскрипции выделяют три стадии: инициацию, элонгацию и терминацию. На стадии инициации РНК-полимераза, взаимодействуя с промотором, вызывает расхождение цепей ДНК и начинает синтез молекулы РНК. В ходе элонгации новосинтезированная цепь РНК образует (за счет спаривания ее оснований с основаниями матричной цепи ДНК) короткие отрезки гибридной двойной спирали ДНК – РНК, которые необходимы для правильного считывания цепи ДНК. Как только РНК-полимераза достигнет терминирующих последовательностей, запускается последняя стадия – терминация. По ее завершении происходит освобождение вновь синтезированной цепи РНК (рис. 48).
Рис. 48. Стадии транскрипции
Синтезированные молекулы РНК могут подвергаться посттранскрипционным ковалентным модификациям, так называемому процессингу (созреванию). Наиболее ярко процессинг выражен у эукариот.
Процессинг РНК
Рассмотрим процессинг иРНК эукариот.
иРНК синтезируется в виде предшественника пре-иРНК. Модификация пре-иРНК начинается на стадии элонгации, в это время происходит кэпирование 5’-конца. Остаток ГТФ присоединяется своим 5’-концом к 5’-концу пре-иРНК с образованием так называемого кэпа. После завершения транскрипции к 3’-концу присоединяется полиА-последовательность, состоящая из 100 – 200 последовательно соединенных адениновых нуклеотидов. В составе пре-иРНК могут содержаться интроны и экзоны. Интроны при созревании РНК вырезаются, концы экзонов соединяются друг с другом. Этот процесс называется сплайсингом. В результате сплайсинга зрелая молекула иРНК становится приблизительно в 4 раза короче пре-иРНК. Этапы процессинга иРНК представлены на рис. 49. Процессинг для иРНК прокариот не характерен.
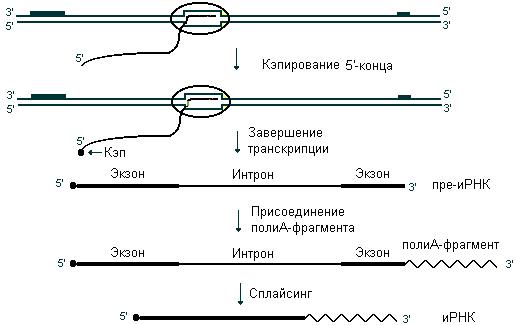
Рис. 49. Процессинг иРНК
рРНК и тРНК также синтезируются в виде более длинных предшественников, которые затем расщепляются и модифицируются. У прокариот рРНК представлены тремя молекулами: 16S-рРНК, 23S-рРНК и 5S-рРНК, у эукариот – четырьмя: 18S-рРНК, 28S-рРНК, 5,8S-рРНК и 5S-рРНК. рРНК как прокариот, так и эукариот образуются из предшественников – пре-рРНК. Предшественник расщепляется, образуя индивидуальные рРНК (рис. 50). рРНК и тРНК не содержат кэпа и полиА-последовательности. У эукариот рРНК и тРНК могут подвергаться сплайсингу. В процессе созревания у некоторых предшественников тРНК наряду с удалением концевых последовательностей может происходить и присоединение нуклеотидных последовательностей, играющих важную роль в их функционировании.
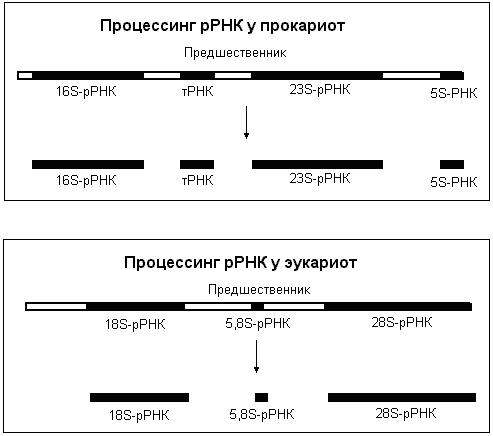
Рис.50. Процессинг рРНК.
Обратная транскрипция
Синтез ДНК при использовании в качестве матрицы РНК носит название обратной транскрипции. Данный процесс катализирует фермент обратная транскриптаза или ревертаза. Существование обратных транскриптаз в составе РНК-содержащих вирусов было показано Г.Темином и Д. Балтимором. Обнаружение обратной транскриптазы позволило ответить на вопрос: как генетическая информация РНК-содержащих вирусов может включиться в ДНК клетки-хозяина. Процесс обратной транскрипции, катализируемый ревертазой, и последующая интеграция генетического материала в геном клетки хозяина представлены на рис. 51. В процессе обратной транскрипции вначале образуется дуплекс РНК – ДНК, затем РНК в составе этого дуплекса разрушается, синтезированная цепь ДНК далее служит матрицей для синтеза второй цепи ДНК. На заключительной стадии происходит интеграция ДНК, синтезированной в результате обратной транскрипции, в ДНК клетки-хозяина. Обратные транскриптазы способны синтезировать ДНК, комплементарную самым различным РНК. Благодаря этой особенности ревертаза нашла широкое применение в научных исследованиях. С помощью обратной транскриптазы можно получить, например, искусственный ген, используя в качестве матрицы иРНК.
Источник
Биология. 10 класс
Генетическая информация в клетке
Хранение, передача и реализация наследственной информации в клетке. Ген. Геном. Реакции матричного синтеза
Необходимо запомнить
На Земле живёт около 7 млрд людей. Если не считать 25–30 млн пар однояйцовых близнецов, то генетически все люди разные: каждый уникален, обладает неповторимыми наследственными особенностями, свойствами характера, способностями, темпераментом.
Такие различия объясняются различиями в генотипах – наборах генов организма; у каждого он уникален. Генетические признаки конкретного организма воплощаются в белках – следовательно, и строение белка одного человека отличается, хотя и совсем немного, от белка другого человека.
Это не означает, что у людей не встречается совершенно одинаковых белков. Белки, выполняющие одни и те же функции, могут быть одинаковыми или совсем незначительно отличаться одной-двумя аминокислотами друг от друга. Но не существует на Земле людей (за исключением однояйцовых близнецов), у которых все белки были бы одинаковы.
Информация о первичной структуре белка закодирована в виде последовательности нуклеотидов в участке молекулы ДНК – гене – единице наследственной информации организма. Каждая молекула ДНК содержит множество генов. Совокупность всех генов организма составляет его генотип.
Кодирование наследственной информации происходит с помощью генетического кода, который универсален для всех организмов. Каждая аминокислота кодируется тремя нуклеотидами (триплетом) ДНК, комбинирующихся в разной последовательности (ААТ, ГЦА, АЦГ, ТГЦ и т.д. Аминокислот, входящих в состав белков – 20, а возможностей для комбинаций четырёх нуклеотидов в группы по три – 64, поэтому одна аминокислота может кодироваться несколькими триплетами. Часть триплетов вовсе не кодирует аминокислоты, а запускает или останавливает биосинтез белка.
ДНК непосредственно не принимает участия в биосинтезе белка. Информация с ДНК сначала копируется на иРНК (транскрипция), а затем на рибосомах переводится в последовательность аминокислот в молекулах синтезируемого белка (процесс трансляции).
В состав и-РНК входят нуклеотиды АЦГУ, триплеты которых называются кодонами: кодоны иРНК комплементарны триплетам ДНК: триплет на ДНК ЦГТ на и-РНК станет триплетом ГЦА, а триплет ДНК ААГ станет триплетом УУЦ.
Таким образом, генетический код – единая система записи наследственной информации в молекулах нуклеиновых кислот в виде последовательности нуклеотидов. Генетический код основан на использовании алфавита, состоящего всего из четырёх букв-нуклеотидов, отличающихся азотистыми основаниями: А, Т (У), Г, Ц.
Основные свойства генетического кода:
1. Генетический код триплетён. Триплет (кодон) – последовательность трёх нуклеотидов, кодирующая одну аминокислоту. Поскольку в состав белков входит 20 аминокислот, то очевидно, что каждая из них не может кодироваться одним нуклеотидом. Двух нуклеотидов для кодирования аминокислот также не хватает, поскольку в этом случае могут быть закодированы только 16 аминокислот. Значит, наименьшее число нуклеотидов, кодирующих одну аминокислоту, оказывается равным трём. (В этом случае число возможных триплетов нуклеотидов составляет 43 = 64).
2. Избыточность (вырожденность) кода является следствием его триплетности и означает то, что одна аминокислота может кодироваться несколькими триплетами (поскольку аминокислот 20, а триплетов – 64), за исключением метионина и триптофана, которые кодируются только одним триплетом. Кроме того, некоторые триплеты выполняют специфические функции: в молекуле иРНК триплеты УАА, УАГ, УГА – являются терминирующими кодонами, т. е. стоп-сигналами, прекращающими синтез полипептидной цепи. Триплет, соответствующий метионину (АУГ), стоящий в начале цепи ДНК, не кодирует аминокислоту, а выполняет функцию инициирования (начала) синтеза.
3. Одновременно с избыточностью коду присуще свойство однозначности: каждому кодону соответствует только одна определённая аминокислота.
4. Код коллинеарен, т. е. последовательность нуклеотидов в гене точно соответствует последовательности аминокислот в белке.
5. Генетический код неперекрываем. Это значит, что процесс считывания не допускает возможности перекрывания кодонов (триплетов).
6. Генетический код универсален, т. е. одинаковые для всех живых организмов вне зависимости от уровня организации и систематического положения этих организмов.
7. Генетический код содержит «знаки препинания» – стоп-кодоны. Начавшись на определённом кодоне, считывание идёт непрерывно триплет за триплетом вплоть до стоп-сигналов (терминирующих кодонов).
Источник
Вопрос 47. Способы передачи генетической информации
Быстрая реакция на изменения условий окружающей среды и приобретение необходимых генетических признаков (напр. изменение вирулентных свойств, устойчивость к АМП). Обмен генетической информацией (ОГИ)- механизм, обеспечивающий многообразие микробного мира.
ОГИ способствует быстрой адаптации бактерий.
ОГИ приводит к образованию рекомбинантной ДНК.
ОГИ служит важным фактором эволюции бактерий.
3 способа передачи генетической информации:
1. Конъюгация – внедрение ДНК из бактерии – донора в клетку реципиента
2. Трансформация – поглощение свободной ДНК из внешней среды
3. Трансдукция – распространение генов бактерий умеренными фагами
Конъюгация
Конъюгация – однонаправленный перенос генетического материала (хромосомной и плазмидной ДНК) от донора к реципиенту при непосредственном контакте клеток.
Донор – бактерия, содержащая конъюгативную плазмиду или конъюгативный транспозон, способная передавать ДНК реципиенту путем горизонтального переноса генов.
Реципиент – любая бактериальная клетка, способная принимать ДНК от бактерии-донора путем горизонтального переноса генов.
Механизм передачи конъюгативных плазмид между Гр- бактериями:
1. Образование межклеточного контакта — конъюгативных пилей
2. Сайт-специфическое разрезание одной цепочки ДНК
3. Сборка особой структуры — релаксосомы для переноса ДНК
4. Перемещение разделенных цепей ДНК в клетку реципиента
5. Синтез комплементарных цепей ДНК
6. Терминация переноса ДНК и разделение конъюгирующих клеток
Трансформация
Трансформация —поглощение бактерией ДНК из окружающей среды и включение в свой геном, при этом бактерия становится генетически трансформированной.
Впервые этот феномен описан у Streptococcus pneumoniae, когда непатогенные живые мутантные клетки стали патогенными для мышей при одновременной инъекции с убитыми нагреванием вирулентными клетками S.pneumoniae.
Изменение происходило в результате трансформирующего действия ДНК из убитых нагреванием вирулентных клеток S.pneumoniae.
Схема трансформации
1. Адсорбция ДНК на клетке (клеточной стенке)
2. Проникновение внутрь клетки
3. Рекомбинация с ДНК клеточной хромосомы
Трансдукция
Трансдукция — обмен генетическим материалом с участием бактериофагов.
Бактериофаги – вирусы бактерий — молекулярные паразиты, использующие для самовоспроизводства аппарат репликации ДНК и синтеза белка зараженных ими клеток.
Трансдукцию осуществляют бактериофаги, содержащие невирусную генетическую информацию. Подобно плазмидам, бактериофаги могут выступать в качестве векторов переноса ДНК от клеток-хозяев. Перенос фаговой информации может служить механизмом обмена генетическим материалом между бактериями. Бактериофаги E.coli
Виды трансдукции:
1. Специфическая трансдукция – перенос генов клетки-хозяина из строго определенных участков хромосомы бактериальной клетки. (Например, 5 из 11 фагов возбудителя дифтерии Corynebacterium diphtheriae – переносчики генов дифтерийного токсина).
2.Неспецифическая трансдукция (общая) — случайный перенос различных генов клетки-хозяина, расположенных в разных участках генома бактерии..Схема трансдукции 
Вопрос 48. Размножение бактерий (клеточный цикл)
У всех живых организмов рост клеток – это увеличение массы и последующее деление с образованием двух идентичных клеток. Не являются исключением и бактерии.
У бактерий генетический материал — в ковалентно замкнутой кольцевой молекуле ДНК.
Молекула ДНК – хромосома. Хромосома расположена в нуклеоиде.
Деление клетки Neisseria gonorrhoeae
Период от деления до деления называется вегетативным клеточным циклом (ВКЦ), он включает несколько этапов:
Репликация ДНК – удвоение генетического материала
Расхождение двух наборов хромосом
Деление клетки
Особенности клеточного цикла прокариот: ВКЦ прокариот и эукариот во многом сходен. Однако есть отличие: Во время быстрого роста в одной бактериальной клетке может происходить 2 — 3 цикла репликации хромосом одновременно.
Источник



