Сила — векторная величина, являющаяся мерой механического воздействия на материальную точку или тело со стороны других тел или полей (Б. М. Яворский, А. А. Детлаф, 1974). Сила полностью задана, если указаны ее численное значение, направление и точка приложения.
В теории и методике физического воспитания рассматривают физическое качество силу как способность человека напряжением мышц преодолевать механические и биомеханические силы, препятствующие действию (Зациорский В. М., 1972; Л. П. Матвеев, 1991).
Мышцы могут проявлять силу: без изменения своей длины (изометрический режим), при уменьшении длины (изотонический режим), при удлинении (эксцентрический режим), при использовании специальной аппаратуры возможно соблюдение изокинетического режима (в ходе сокращения мышц соблюдается либо постоянная скорость, либо сила).
Силовое проявление мышцы зависит от:
— интенсивности активации мотнейронного пула спинного мозга данной мышцы;
— количества активированных двигательных единиц и мышечных волокон;
— количества миофибрилл в каждом мышечном волокне;
— скорости сокращения миофибрилл, которая зависти от активности миозиновой АТФ-азы и величины внешнего сопротивления;
— законов механики мышечного сокращения (сила — длина мышцы, сила — скорость сокращения);
— начального состояния исполнительного аппарата (утомленные мышечные волокна демонстрируют меньшую силу).
Спортсмен при желании сократить какую-либо мышцу активизирует соответствующий двигательный нейрон в коре головного мозга, который посылает импульсы в спинной мозг к мотонейронному пулу, обслуживающему данную мышцу. Поскольку в мотонейронном пуле размеры мотонейронов различаются, то при низкой частоте импульсации из ЦНС могут активироваться только низкопороговые мотонейроны. Каждый мотонейрон иннервирует свои мышечные волокна. Поэтому активация мотонейрона приводит к рекрутированию или возбуждению соответствующих мышечных волокон. Каждое активное мышечное волокно под влиянием электрических импульсов выпускает из СПР ионы кальция, которые снимают ингибитор с активных центров актина. Это обеспечивает образование актин-миозиновых мостиков и начало их поворота и мышечного сокращения. На поворот мостиков и отсоединение актина от миозина тратится энергия одной молекулы АТФ. Продолжительность работы мостика составляет 1 мс. Вероятность образования мостиков зависит от взаимного расположения между собой нитей актина и миозина, отсюда возникает зависимость сила — длина активной мышца, а также от скорости взаимного перемещения (скольжения) их одной по отношению к другой, соответственно, имеем зависимость «сила-скорость».
Зависимость «сила — длина активного мышечного волокна» определяется, как правило, относительным расположением между собой головок миозина и активных центров актина. Максимальное количество мостиков возникает при некоторой средней длине мышцы. Отклонение от этой длины в большую или меньшую сторону ведет к снижению силовых проявлений мышечного волокна (мышцы). Однако, в случае растяжения некоторых мышц, еще не в активном состоянии, могут возникать значительные силы сопротивления растяжению, например, в мышцах сгибателях голеностопного или лучезапястного сустава. Эти силы связаны с растяжением соединительных тканей, например, перемезиума. В биомеханике в таком случае говорят о параллельном упругом компоненте мышцы. Упругостью обладают сухожилия, зет-пластинки саркомеров и нити миозина, к которым прикреплены головки. Такую упругость называют последовательной упругой компонентой.
Наличие последовательной упругой компоненты в мышечных волокнах приводит к тому, что с ростом числа рекрутированных МВ увеличивается жесткость мышцы -коэффициент упругости (В. М. Зациорский с соав., 1981).
Растягивание активной мышцы приводит не только к накоплению энергии упругой деформации в последовательной упругой компоненте, но и к прекращению работы мостиков, а именно, они перестают отцепляться за счет энергии молекул АТФ. Разрыв мостиков происходит благодаря действию внешней — механической силы. В итоге отрицательная работа мышц выполняется с очень высоким коэффициентов полезного действия, с минимальными затратами АТФ, а значит и кислорода.
Источник
Сокращение скелетной мышцы
Основными физиологическими свойствами мышц являются их возбудимость, проводимость и сократимость. Последняя проявляется или в укорочении мышцы, или в развитии напряжения.
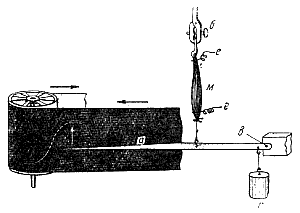
Миография . Для регистрации мышечного сокращения применяется методика миографии, т. е. графической регистрации сокращения скелетных мышц с помощью рычажка, присоединенного к одному концу мышцы ( рис. 139 ). Свободный конец рычажка чертит на ленте кимографа кривую сокращения- миограмму. Этот способ регистрации мышечного сокращения прост и не требует сложного оборудования, но имеет тот недостаток, что инерция рычага и его трение по поверхности ленты кимографа несколько искажают запись. Во избежание этого недостатка теперь применяют специальный датчик, преобразующий механические изменения (линейные перемещения или усилия мышцы) в колебания силы электрического тока. Последние регистрируются с помощью шлейфного или катодного осциллографа.
Точной методикой является также оптическая регистрация, производимая на фотокимографе с помощью пучка света, отраженного от зеркальца, наклеенного на брюшко мышцы.
Изотоническое и изометрическое сокращения . Изотоническим сокращением называется такое сокращение мышцы, при котором ее волокна укорачиваются, но напряжение остается постоянным. Примером изотонического сокращения является свободное поднятие мышцей груза, который обусловливает ее постоянное напряжение. Изометрическим называется такое сокращение, при котором мышца укоротиться не может, например если оба конца ее неподвижно закреплены.
В этом случае длина мышечных волокон остается неизменной, но зато напряжение их возрастает. Для записи изометрического сокращения мышцу заставляют работать, прикрепив к ней тугую пружину, которая почти не позволяет ей укоротиться. В подобном случае, пользуясь различного рода усилителями, записывают то ничтожное (порядка нескольких микронов) укорочение мышцы, которое, в сущности говоря, отображает развитие ее напряжения.
Естественные сокращения мышц в организме никогда не бывают чисто изотоническими или чисто изометрическими, так как мышцы, поднимая груз (например, сгибая конечность в суставе), укорачиваются и вместе с тем, растягивая антагонистические мышцы, меняют свое напряжение.
Методы раздражения мышц . Для того чтобы в эксперименте вызвать сокращение мышцы, ее подвергают раздражению. Непосредственное раздражение самой мышцы (например, электрическим током) называется прямым раздражением; раздражение двигательного нерва, ведущее к сокращению интернированной этим нервом мышцы, называется непрямым раздражением. Ввиду того что возбудимость мышечной ткани меньше, чем нервной, приложение электродов раздражающего тока непосредственно к мышце еще не обеспечивает прямого раздражения: ток, распространяясь по мышечной ткани, действует в первую очередь на находящиеся в ней окончания двигательных нервов и возбуждает их, что ведет к сокращению мышцы.
Чтобы получить сокращение мышцы под влиянием прямого раздражения, необходимо либо выключить в ней двигательные нервные окончания ядом кураре, либо прикладывать стимул через введенный внутрь мышечного волокна микроэлектрод.
Рис. 139. Методика миографии для регистрации изотонического сокращения мышцы (по И. С. Беритову). М — мышца, при сокращении поднимающая рычаг а и груз г, привязанный у оси рычага (в); б — мышечная клемма; е — электроды для раздражения.
Источник
Суммация сокращений и тетанус
Если в эксперименте на отдельное мышечное волокно или на всю мышцу действуют два быстро следующих друг за другом сильных одиночных раздражения, то возникающее сокращение будет иметь большую амнли гуду, чем максимальное сокращение п.р:н, одиночном раздражении. Сократительные эффекты, вызванные первым и вторым раздражениями, как бы складываются. Это явление называется суммацией сокращений (рис. 29). Оно наблюдается как при прямом, так и при непрямом раздражении мышцы. Для возникновения суммации необходимо, чтобы интервал между раздражением имел определенную длительность: он должен быть длиннее рефракторного периода, иначе на второе раздражение не будет ответа, и короче всей длительности сократительного ответа, чтобы второе раздражение подействовало на мышцу раньше, чем она успеет расслабиться после первого раздражения. При этом возможны два варианта. Если второе раздражение поступает, когда мышца уже начала расслабляться, то на мио-
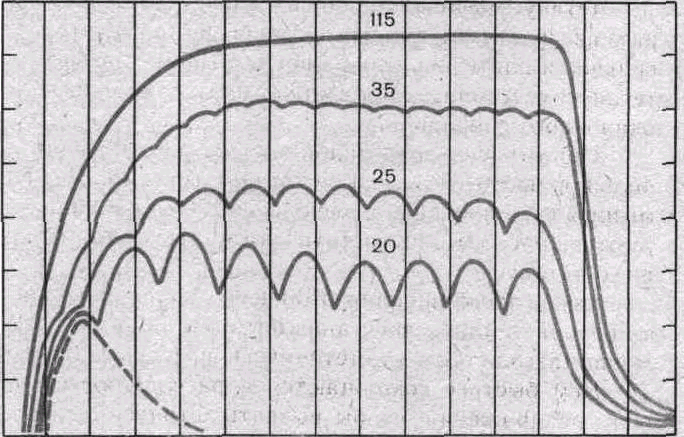
Рис. 30. Сокращения икроножной мышцы при разной частоте раздражения (по Е. F. Жукову).
Одиночное сокращение показано пунктиром. Цифры над кривыми — частоты стимулов. При частотах 20—35 в секунду — зубчатый тетанус, при частоте 115 в секунду— гладкий тетанус.
-Рис. 31. Тетанус изолированного мышечного волокна (по Ф. Н. Серкову).
а — зубчатый тетанус при частоте раздражения 18 Гц; б — гладкий тетанус при частоте раздражения 35 Гц; М — миограмма; Р — отметки раздражения; В — отметка времени 1 с.
Рис. 32. Одновременная запись сокращения (а) и электрической активности (б) скелетной мышцы кошки при тетаническом раздражении нерва.
графической кривой вершина этого сокращения будет отделена от вершины первого западением (рис. 29, Ж — Г). Если же второе раздражение действует, когда первое еще не дошло до своей вершины, то второе сокращение полностью сливается С первым, образуя единую суммированную вершину (рис. 29, А—В).
Рассмотрим суммацию в икроножной мышце лягушки. Продолжительность восходящей фазы ее сокращения примерно 0,05 с. Поэтому для воспроизведения на этой мышце первого типа суммации сокращений (неполная суммация) необходимо, чтобы интервал между первым и вторым раздражениями был больше 0,05 с, а для получения второго типа суммации (так называемая полная суммация) — меньше 0,05 с.
Как при полной, так и при неполной суммации сокращений потенциалы действия не суммируются.
Тетанус мышцы. Если на отдельное мышечное волокно или на всю мышцу действуют ритмические раздражения с такой частотой, что их эффекты суммируются, наступает сильное и длительное сокращение мышцы, называемое тетаническим сокращением, или тетанусом. Амплитуда его может быть. в несколько раз больше величины максимального одиночного сокращения.
При относительно малой частоте раздражении наблюдается зубчатый тетанус, при большой частоте — гладкий тетанус (рис. 30, 31). При тетанусе сократительные ответы мышцы суммированы, а электрические ее реакции — потенциалы действия — не суммируются (рис. 32) и их частота соответствует частоте ритмического раздражения, вызвавшего тетанус.
После прекращения тетанического раздражения волокна полностью расслабляются, их исходная длина восстанавливается лишь по истечении некоторого времени. Это явление называется послететанической, или остаточной, контрактурой.
Чем быстрее сокращаются и расслабляются волокна мышцы, тем чаще должны быть раздражения, чтобы вызвать тетанус.
Источник
Механизм мышечного сокращения
Содержание
Источник: «Программы тренировок», научное изд. Автор: профессор, доктор наук Тудор Бомпа, 2016 г.
Структура мышц Править
Мышца — это комплексная структура, отвечающая за движение. Мышцы состоят из саркомеров, которые содержат определенное сочетание фибриллярных белков — миозина (толстые нити) и актина (тонкие нити), которые играют важную роль в мышечных сокращениях. Таким образом, саркомер — это сократительный элемент мышечного волокна, состоящий из миозиновых и актиновых белковых нитей.
Помимо этого, способность мышцы сокращаться и прилагать силу зависит конкретно от ее вида, площади поперечного сечения, а также длины и количества волокон внутри мышцы. Число волокон определяется генетикой, и на него невозможно повлиять с помощью тренировок; однако тренировки в состоянии изменить другие переменные. Например, число и толщина миозиновых нитей увеличивается посредством упорных тренировок с максимальной силовой нагрузкой. Увеличение толщины мышечных нитей увеличивает размер мышцы и силу сокращений.
Человеческое тело состоит из различных типов мышечных волокон, подразделяющихся на группы, и каждая группа относится к одной двигательной единице. В общем и целом в нашем организме имеются тысячи двигательных единиц, в которых находятся десятки тысяч мышечных волокон. Каждая двигательная единица содержит сотни или тысячи мышечных волокон, пребывающих в покое до тех пор, пока им не нужно действовать. Двигательная единица управляет совокупностью волокон и направляет их действия по закону «все или ничего». Этот закон означает, что при раздражении двигательной единицы импульс, направляемый в ее мышечные волокна, либо распространяется полностью — таким образом раздражая всю совокупность волокон, — либо не распространяется вообще.
Разные двигательные единицы реагируют на разные нагрузки при тренировках. Например, выполнение жима лежа с 60% повторного максимума задействует определенную совокупность двигательных единиц, тогда как более крупные двигательные единицы ожидают более высокой нагрузки. Поскольку последовательное задействование двигательных единиц зависит от нагрузки, необходимо разрабатывать специальные программы, чтобы активизировать и адаптировать основные группы двигательных единиц и мышечных волокон, играющих доминирующую роль в избранном виде спорта. К примеру, в тренировках для спринта на короткую дистанцию и легкоатлетических дисциплин (таких как толкание ядра) следует использовать тяжелые нагрузки, чтобы способствовать развитию силы, необходимой для оптимизации скорости и взрывных действий.
Мышечные волокна выполняют разные биохимические (метаболические) функции; выражаясь конкретнее, одни лучше приспособлены с физиологической точки зрения к работе в анаэробных условиях, а другие лучше работают в аэробных условиях. Волокна, которые используют кислород для выработки энергии, называются аэробными, тип I, красными или медленными. Волокна, которым кислород не требуется, называются анаэробными, тип II, белыми или быстрыми. Быстрые мышечные волокна, в свою очередь, делятся на подтипы IIА и IIХ (иногда называемые IIВ, хотя у людей тип IIВ практически не встречается [1] ).
Медленные и быстрые волокна существуют примерно в равной пропорции. Однако в зависимости от их функций, в некоторых группах мышц (например, подколенные сухожилия, бицепсы) содержится больше быстрых волокон, тогда как в других (например, в камбаловидной мышце) содержится больше медленных волокон. В таблице 2.1 мы сравниваем характеристики быстрых и медленных волокон.
Сравнение быстрых и медленных волокон
Красные, тип I, аэробные
Белые, тип II, анаэробные
• Нервная клетка меньше — иннервирует от 10 до 180 мышечных волокон
• Развивают долгие, продолжительные сокращения
• Применяются для развития выносливости
• Активизируются во время низко- и высокоинтенсивной деятельности
• Большая нервная клетка — иннервирует
от 300 до 500 (или более) мышечных волокон
• Развивают короткие, сильные сокращения
• Применяются для развития скорости и силы
• Активизируются только во время высокоинтенсивной деятельности
Тренировки могут влиять на эти характеристики. Датские ученые Андерсен и Аагаард [2] [3] [4] [5] [6] в своих исследованиях показывают, что при объемных нагрузках или лактатных по природе тренировках волокна IIХ приобретают характеристики волокон IIА. То есть богатая миозином цепочка этих волокон становится более медленной и более эффективно справляется с лактатной деятельностью. Эти изменения можно повернуть вспять, снижая тренировочную нагрузку (тейперинг), в результате чего волокна IIХ возвращаются к изначальным характеристикам наиболее быстрых волокон [3] . Силовые тренировки также увеличивают размер волокон, благодаря чему вырабатывается больше силы.
Сокращение быстрой двигательной единицы более быстрое и мощное, чем сокращение медленной двигательной единицы. В результате пропорция быстрых волокон, как правило, выше в организме успешных спортсменов, занимающихся скоростно-силовыми видами спорта, но они также быстрее утомляются. Спортсмены с более высоким скоплением медленных волокон, напротив, обычно преуспевают в видах спорта на выносливость, поскольку они могут выполнять нагрузки низкой интенсивности в течение более продолжительного времени.
Активизация мышечных волокон происходит по принципу величины, известному также как принцип Хеннемана [7] , согласно которому двигательные единицы и мышечные волокна активизируются начиная с меньшей в сторону большей. Активация всегда начинается с медленных волокон. При низкой или умеренно интенсивной нагрузке активируются медленные волокна и выполняют большую часть работы. При сильной нагрузке сначала сокращаются медленные волокна, затем в процесс вовлекаются быстрые волокна. При повторениях до отказа с умеренной нагрузкой двигательные единицы, состоящие из быстрых волокон, постепенно активизируются, чтобы поддерживать выработку силы, тогда как ранее задействованные двигательные единицы утомляются (см. рис. 1).
В распределении типов мышечных волокон у спортсменов, занимающихся разными видами спорта, могут наблюдаться различия. Это иллюстрируют рис. 2 и 2.3, представляющие общий процент содержания быстрых и медленных мышечных волокон у спортсменов в избранных видах спорта. Например, существенная разница между спринтерами и марафонцами четко дает понять, что успех в некоторых видах спорта хотя бы частично определяется генетическим составом мышечных волокон спортсмена.
Следовательно, пиковая мощность, вырабатываемая спортсменами, также имеет отношение к распределению типов волокон — чем выше процент быстрых волокон, тем большую мощность развивает спортсмен. Процент быстрых волокон также имеет отношение к скорости: чем выше скорость спортсмена, тем выше процент имеющихся у него быстрых волокон. Из таких людей получаются превосходные спринтеры и прыгуны, а подобный природный талант следует направлять в русло скоростно-силовых видов спорта. Попытка тренировать их, скажем, для бега на дистанцию означает трату таланта; в таких дисциплинах их ждет лишь средний успех, тогда как из них могут выйти отличные спринтеры, бейсболисты или футболисты (на этом список скоростносиловых видов спорта не кончается).
Как мы описывали раньше, мышечные сокращения происходят в результате цепочки событий с участием белковых нитей — миозина и актина. В миозиновых нитях содержатся поперечные мостики — крошечные перемычки, выступающие вбок по направлению к актиновым нитям. Возбуждение, приводящее к сокращениям, стимулирует все волокно, создавая химические изменения, позволяющие актиновым нитям соединяться с миозиновыми поперечными мостиками. Связывание миозина с актином посредством поперечных мостиков высвобождает энергию, из-за чего поперечные мостики поворачиваются, таким образом подтягивая или совершая скользящее движение, связывающее миозиновые нити с актиновыми. Это скользящее движение вызывает мышечное сокращение, которое вырабатывает силу.
Чтобы визуализировать это иначе, вообразите гребную лодку. Весла представляют собой миозиновые нити, а воды — актиновые. Когда весла ударяются о воду, лодка с силой тянется вперед — и чем больше в воде весел, чем выше физическая сила гребца, тем больше вырабатываемая сила. Увеличение количества и толщины миозиновых нитей таким же образом повышает выработку силы.
Описанная ранее теория скользящих нитей дает понять, как работают мышцы, чтобы выработать силу. Эта теория включает в себя механизмы, способствующие эффективным мышечным сокращениям. Например, освобождение запаса эластичной энергии и рефлекторная адаптация играют ключевую роль в оптимизации спортивной работоспособности, но подобная адаптация происходит только тогда, когда в процессе тренировки происходит правильная стимуляция. Например, способность спортсмена использовать запас энергии для того, чтобы прыгать выше или толкать ядро дальше, оптимизируется посредством взрывных движений, как те, которые используются в плиометрическом тренинге. Однако мышечные компоненты — как, например, эластичные компоненты (сюда входят сухожилия, мышечные волокна и поперечные мостики) — не могут осуществлять эффективную транспортировку энергии, если спортсмен не укрепляет параллельные эластичные компоненты (напр., связки) и коллагеновые структуры (обеспечивающие стабильность и предохраняющие от травм). Если телу нужно выдерживать силы и воздействия, которым спортсмен подвергается, чтобы оптимизировать эластичные качества мышц, анатомическая адаптация должна предшествовать силовому тренингу.
Рефлекс — это непроизвольное мышечное сокращение, вызванное внешним стимулом [8] . Два основных компонента контроля рефлексов — это мышечные веретена и нервносухожильное веретено. Мышечные веретена реагируют на величину и скорость мышечного растяжения [9] , тогда как нервно-сухожильное веретено (которое находится в местах соединения мышечных волокон с сухожильными пучками [8] ) реагирует на мышечное напряжение. Когда в мышцах развивается высокая степень напряжения или растяжения, мышечные веретена и нервно-сухожильное веретено непроизвольно расслабляют мышцу, чтобы защитить ее от повреждения и травмы.
При пресечении этих ингибиторных реакций повышается спортивная работоспособность. Единственный способ добиться этого — адаптировать организм к более высокой степени напряжения, что повышает порог активизации рефлексов. Этой адаптации можно добиться посредством силового тренинга с использованием постепенно утяжеляющейся нагрузки (до 90 процентов повторного максимума или даже выше), таким образом вынуждая нервно-мышечную систему выдерживать более высокое напряжение, постоянно задействуя большее число быстрых волокон. В быстрых волокнах вырабатывается больше белка, что способствует увеличению силы.
Все спортивные движения выполняются по двигательной модели, которая называется циклом растяжение — сокращение и характеризуется тремя основными типами сокращения: эксцентрическим (удлинение), изометрическим (статичное положение) и концентрическим (сокращение). Например, волейболист, который быстро приседает и сразу подпрыгивает, чтобы блокировать атакующий удар, выполнил весь цикл растяжение — сокращение. То же касается и спортсмена, который опускает штангу на грудь и быстро выполняет взрывное движение, вытягивая руки. Чтобы полноценно пользоваться физиологическими качествами цикла растяжение — сокращение, мышца должна быстро переходить от удлинения к сокращению [10] (Schmidtble-icher, 1992).
Мышечный потенциал оптимизируется, когда активизируются все сложные факторы, влияющие на цикл растяжение — сокращение. Их влияние можно использовать для улучшения спортивных показателей только тогда, когда нервно-мышечная система стратегически стимулируется в правильной последовательности. Именно для достижения этой цели периодизация тренировки силы основывает планирование этапов на физиологической базе выбранного вида спорта. После составления эргогенного профиля (оценки вклада энергетических систем) выбранного вида спорта нужно пошагово распланировать этапы тренировки, чтобы перенести положительную нервно-мышечную адаптацию на практические показатели деятельности человека. Таким образом, понимание прикладной человеческой физиологии и установление цели в конце каждого этапа помогают тренерам и спортсменам интегрировать физиологические принципы в конкретную спортивную тренировку.
Повторим: скелетно-мышечная система тела — это сочетание костей, прикрепляемых друг к другу с помощью связок в области суставов. Пересекающие эти суставы мышцы дают силу для движения тела. Однако скелетные мышцы не сокращаются независимо друг от друга. Движения, выполняемые вокруг сустава, производятся несколькими мышцами, каждая из которых выполняет определенную роль, как уже было упомянуто выше.
Агонисты — или синергисты — это мышцы, которые взаимодействуют друг с другом при выполнении движения. В большинстве случаев, особенно если речь идет об умелом и опытном спортсмене, мышцы-антагонисты расслабляются, облегчая движение. Поскольку взаимодействие мышц группы агонистов и антагонистов напрямую влияет на спортивные движения, неправильное взаимодействие между этими группами может привести к порывистому или скованному движению. Следовательно, гладкость мышечного сокращения можно улучшить, если сосредоточиться на расслаблении антагонистов.
По этой причине одновременное сокращение (одновременная активизация мышц-агонистов и антагонистов, чтобы стабилизировать сустав) рекомендуется только на ранних стадиях реабилитации после травмы. Здоровому же спортсмену, особенно если он занимается силовыми видами спорта, не нужно выполнять упражнения (например, на нестабильной поверхности), вызывающие одновременные сокращения. К примеру, одной из основных характеристик элитных спринтеров является очень низкая миоэлектрическая активность мышц-антагонистов в каждой фазе цикла шага [11] .
Первичные мышцы в первую очередь отвечают за суставное действие, которое является частью объемного силового движения или технической способности. Например, во время флексии локтя (сгибание бицепса) первичной мышцей является двуглавая мышца, тогда как трехглавая мышца (трицепс) выступает в роли антагониста и должна быть расслаблена, чтобы обеспечить беспрепятственное действие. В дополнение к этому стабилизаторы, или фиксаторы (обычно это меньшие мышцы), сокращаются изометрически, чтобы закрепить кость так, чтобы у первичных мышц была прочная база, откуда начинать натяжение. Мышцы других конечностей также могут принимать в этом участие, выступая в роли стабилизаторов, позволяющих первичным мышцам выполнять необходимые движения. Например, когда дзюдоист тянет соперника на себя, удерживая его за дзюдоги, мышцы его спины, ног и живота сокращаются изометрически, чтобы обеспечить стабильное основание для действия локтевых сгибателей (бицепсов), плечевых разгибателей (задние дельты) и лопаточных аддукторов и депрессоров (трапециевидная мышца и широчайшая мышца спины).
Если мышцу стимулировать коротким электрическим импульсом, спустя небольшой латентный период происходит ее сокращение. Такое сокращение называется «одиночное сокращение мышцы». Одиночное мышечное сокращение длится около 10-50 мс, причем оно достигает максимальной силы через 5-30 мс.
Каждое отдельное мышечное волокно подчиняется закону «все или ничего», т. е. при силе раздражения выше порогового уровня происходит полное сокращение с максимальной для данного волокна силой, а ступенчатое повышение силы сокращения по мере увеличения силы раздражения невозможно. Поскольку смешанная мышца состоит из множества волокон с различным уровнем чувствительности к возбуждению, сокращение всей мышцы может быть ступенчатым в зависимости от силы раздражения, при этом при сильных раздражениях происходит активация глубжележащих мышечных волокон.
Механизм скольжения филаментов Править
Укорочение мышцы происходит за счет укорочения образующих ее саркомеров, которые, в свою очередь, укорачиваются за счет скольжения относительно друг друга актиновых и миозиновых филаментов (а не укорочения самих белков). Теория скольжения филаментов была предложена учеными Huxley и Hanson (Huxley, 1974; рис. 1). (В 1954 г. две группы исследователей — X. Хаксли с Дж. Хэнсон и А. Хаксли с Р. Нидергерке — сформулировали теорию, объясняющую мышечное сокращение скольжением нитей. Независимо друг от друга они обнаружили, что длина диска А оставалась постоянной в расслабленном и укороченном саркомере. Это позволило предположить, что есть два набора нитей — актиновые и миозиновые, причем одни входят в промежутки между другими, и при изменении длины саркомера эти нити каким-то образом скользят друг по другу. Сейчас эта гипотеза принята почти всеми.)
Актин и миозин — два сократительных белка, которые способны вступать в химическое взаимодействие, приводящее к изменению их взаимного расположения в мышечной клетке. При этом цепочка миозина прикрепляется к актиновой нити с помощью целого ряда особых «головок», каждая из которых сидит на длинной пружинистой «шее». Когда происходит сцепление между миозиновой головкой и актиновой нитью, конформация комплекса этих двух белков изменяется, миозиновые цепочки продвигаются между актиновыми нитями и мышца в целом укорачивается (сокращается). Однако, чтобы химическая связь между головкой миозина и активной нитью образовалась, необходимо подготовить этот процесс, поскольку в спокойном (расслабленном) состоянии мышцы активные зоны белка актина заняты другим белком — тропохмиозином, который не позволяет актину вступить во взаимодействие с миозином. Именно для того, чтобы убрать тропомиозиновый «чехол» с актиновой нити, требуется быстрое выливание ионов кальция из цистерн саркоплазматического ретикулума, что происходит в результате прохождения через мембрану мышечной клетки потенциала действия. Кальций изменяет конформацию молекулы тропомиозина, в результате чего активные зоны молекулы актина открываются для присоединения головок миозина. Само это присоединение осуществляется с помощью так называемых водородных мостиков, которые очень прочно связывают две белковые молекулы — актин и миозин — и способны в таком связанном виде находиться очень долго.
Для отсоединения миозиновой головки от актина необходимо затратить энергию аденозинтрифосфа-та (АТФ), при этом миозин выступает в роли АТФазы (фермента, расщепляющего АТФ). Расщепление АТФ на аденозиндифосфат (АДФ) и неорганический фосфат (Ф) высвобождает энергию, разрушает связь между актином и миозином и возвращает головку миозина в исходное положение. В дальнейшем между актином и миозином могут снова образовываться поперечные связи.
При отсутствии АТФ актин-миозиновые связи не разрушаются. Это и является причиной трупного окоченения (rigor mortis) после смерти, т. к. останавливается выработка АТФ в организме — АТФ предотвращает мышечную ригидность.
Даже при мышечных сокращениях без видимого укорочения (изометрические сокращения, см. выше) активируется цикл формирования поперечных связей, мышца потребляет АТФ и выделяет тепло. Головка миозина многократно присоединяется на одно и то же место связывания актина, и вся система миофиламентов остается неподвижной.
Внимание: Сократительные элементы мышц актин и миозин сами по себе не способны к укорочению. Мышечное укорочение является следствием взаимного скольжения миофиламентов относительно друг друга (механизм скольжения филаментов).
Как же образование поперечных связей (водородных мостиков) переходит в движение? Одиночный саркомер за один цикл укорачивается приблизительно на 5-10 нм, т.е. примерно на 1 % своей общей длины. За счет быстрого повторения цикла поперечных связей возможно укорочение на 0,4 мкм, или 20% своей длины. Поскольку каждая миофибрилла состоит из множества саркомеров и во всех них одновременно (но не синхронно) образуются поперечные связи, суммарно их работа приводит к видимому укорочению всей мышцы. Передача силы этого укорочения происходит через Z-линии миофибрилл, а также концы сухожилий, прикрепленных к костям, в результате чего и возникает движение в суставах, через которые мышцы реализуют перемещение в пространстве частей тела или продвижение всего тела.
Наибольшую силу сокращений мышечные волокна развивают при длине 2-2,2 мкм. При сильном растяжении или укорочении саркомеров сила сокращений снижается (рис. 2). Эту зависимость можно объяснить механизмом скольжения филаментов: при указанной длине саркомеров наложение миозиновых и актиновых волокон оптимально; при большем укорочении миофиламенты перекрываются слишком сильно, а при растяжении наложение миофиламентов недостаточно для развития достаточной силы сокращений.
Скорость укорочения мышцы зависит от нагрузки на эту мышцу (закон Хилла, рис. 3). Она максимальна без нагрузки, а при максимальной нагрузке практически равна нулю, что соответствует изометрическому сокращению, при котором мышца развивает силу, не изменяя своей длины.
Важным фактором, влияющим на силу сокращений, является величина растяжения мышцы. Тяга за конец мышцы и натяжение мышечных волокон называются пассивным растяжением. Мышца обладает эластическими свойствами, однако в отличие от стальной пружины зависимость напряжения от растяжения не линейна, а образует дугообразную кривую. С увеличением растяжения повышается и напряжение мышцы, но до определенного максимума. Кривая, описывающая эти взаимоотношения, называется кривой растяжения в покое.
Данный физиологический механизм объясняется эластическими элементами мышцы — эластичностью сарколеммы и соединительной ткани, располагающимися параллельно сократительным мышечным волокнам.
Также при растяжении изменяется и наложение друг на друга миофиламентов, однако это не оказывает влияния на кривую растяжения, т. к. в покое не образуются поперечные связи между актином и миозином. Предварительное растяжение (пассивное растяжение) суммируется с силой изометрических сокращений (активная сила сокращений).