Синaпс – специализированный контакт между нервными клетками (или нервными и другими возбудимыми клетками), обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов нервные клетки объединяются в нервные сети, которые осуществляют обработку информации. Взаимосвязь между нервной системой и периферическими органами и тканями также осуществляется при помощи синапсов.
Классификация синапсов
По морфологическому принципу синапсы подразделяют на:
нейро-мышечные (аксон нейрона контактирует с мышечной клеткой);
нейро-секреторные (аксон нейрона контактирует с секреторной клеткой);
нейро-нейрональные (аксон нейрона контактирует с другим нейроном):
аксо-соматические (с телом другого нейрона), аксо-аксональные (с аксоном другого нейрона), аксо-дендритические (с дендритом другого нейрон).
По способу передачи возбуждения синапсы подразделяют на:
электрические (возбуждение передается при помощи электрического тока);
химические (возбуждение передается при помощи химического вещества):
адренергические (возбуждение передается при помощи норадреналина), холинергические (возбуждение передается при помощи ацетилхолина), пептидергические, NO -ергические, пуринергические и т. п.
По физиологическому эффекту синапсы подразделяют на:
возбуждающие (деполяризуют постсинаптическую мембрану и вызывают возбуждение постсинаптической клетки);
тормозные (гиперполяризуют постсинаптическую мембрану и вызывают торможение постсинаптической клетки).
Ультраструктура синапсов
Все синапсы имеют общий план строения (рис. 1).
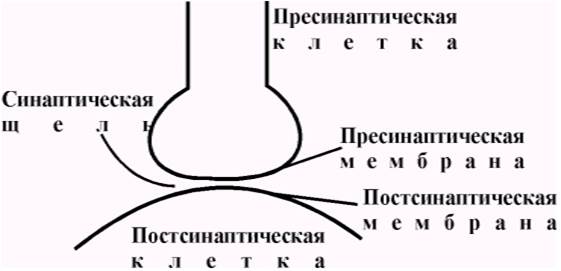
Конечная часть аксона (синаптическое окончание), подходя к иннервируемой клетке, теряет миелиновую оболочку и образует на конце небольшое утолщение (синаптическую бляшку). Ту часть мембраны аксона, которая контактирует с иннервируемой клеткой, называют пресинаптической мембраной. Синаптическая щель – узкое пространство между пресинаптической мембраной и мембраной иннервируемой клетки, которое является непосредственным продолжением межклеточного пространства. Постсинаптическая мембрана – участок мембраны иннервируемой клетки, контактирующий с пресинаптической мембраной через синаптическую щель.
Рис. 1. Ультраструктура химического и электрического синапса.
Особенности ультраструктуры электрического синапса (см. рис. 1):
узкая (около 5 нм) синаптическая щель; наличие поперечных канальцев, соединяющих пресинаптическую и постсинаптическую мембрану.
Особенности ультраструктуры химического синапса (см. рис. 1):
широкая (20–50 нм) синаптическая щель; наличие в синаптической бляшке синаптических пузырьков (везикул), заполненных химическим веществом, при помощи которого передается возбуждение; в постсинаптической мембране имеются многочисленные хемочувствительные каналы (в возбуждающем синапсе – для Nа+ , в тормозном – для Cl – и К +), но отсутствуют потенциалчувствительные каналы.
Механизм передачи возбужденияв электрическом синапсе
Механизм проведения возбуждения аналогичен механизму проведения возбуждения в нервном волокне. Во время развития ПД происходит реверсия заряда пресинаптической мембраны. Электрический ток, возникающий между пресинаптической и постсинаптической мембраной, раздражает постсинаптическую мембрану и вызывает генерацию в ней ПД (рис. 2).
Рис. 2. Передача возбуждения в электрическом синапсе.
Этапы и механизмы передачи возбуждения в возбуждающем химическом синапсе
Передача возбуждения в химическом синапсе – сложный физиологический процесс, протекающий в несколько этапов. На пресинаптической мембране осуществляется трансформация электрического сигнала в химический, который на постсинаптической мембране снова трансформируется в электрический сигнал.
Синтез медиатора
Медиатором (посредником) называют химическое вещество, которое обеспечивает одностороннюю передачу возбуждения в химическом синапсе. Некоторые медиаторы (например, ацетилхолин) синтезируются в цитоплазме синаптического окончания, и там же молекулы медиатора депонируются в синаптических пузырьках. Ферменты, необходимые для синтеза медиатора, образуются в теле нейрона и доставляются в синаптическое окончание путем медленного (1–3 мм/сут) аксонного транспорта. Другие медиаторы (пептиды и др.) синтезируются и упаковываются в везикулы в теле нейрона, готовые синаптические пузырьки доставляются в синаптичекую бляшку за счет быстрого (400 мм/сут) аксонного транспорта. Синтез медиатора и образование синаптических пузырьков осуществляется непрерывно.
Секреция медиатора
Содержимое синаптических пузырьков может выбрасываться в синаптическую щель путем экзоцитоза. При опорожнении одного синаптического пузырька в синаптичекую щель выбрасывается порция (квант) медиатора, которая включает около 10000 молекул.
Для активации экзоцитоза необходимы ионы Са++ . В состоянии покоя уровень Са++ в синаптическом окончании низок и выделения медиатора практически не происходит. Приход в синаптическое окончание возбуждения приводит к деполяризации пресинаптической мембраны и открытию потенциалчувствительных Са++ -каналов. Ионы Са++ поступают в цитоплазму синаптического окончания (рис. 3, А,Б) и активируют опорожнение синаптических пузырьков в синаптическую щель (рис. 3, В).
Рис. 3. Передача сигнала в возбуждающем химическом синапсе. А — Д – последовательность процессов при срабатывании химического синапса; Е – деполяризация постсинаптической мембраны (ВПСП).
Взаимодействие медиатора с рецепторами постсинаптической мембраны
Молекулы медиатора диффундируют через синаптическую щель и достигают постсинаптической мембраны, где связываются с рецепторами хемочувствительных Na+ -каналов (рис. 3, Г). Присоединение медиатора к рецептору приводит к открытию Na+ -каналов, через которые в клетку входят ионы Na+ (рис. 3, Д). В результате входа в клетку положительно заряженных ионов происходит локальная деполяризация постсинаптической мембраны, которую называют возбуждающий постсинаптический потенциал (ВПСП) (рис. 3, Е).
Инактивация медиатора
Ферменты, находящиеся в синаптической щели, разрушают молекулы медиатора. В результате происходит закрытие Na+ -каналов и восстановление МП постсинаптической клетки. Некоторые медиаторы (например, адреналин) не разрушаются ферментами, а удаляются из синаптической щели путем быстрого обратного всасывания (пиноцитоза) в синаптическое окончание.
Генерация ПД
В нейро-мышечном синапсе амплитуда единичного ВПСП достаточно велика. Поэтому для генерации ПД в мышечной клетке достаточно прихода одного нервного импульса. Генерация ПД в мышечной клетке происходит в области, окружающей постсинаптическую мембрану.
В нейро-нейрональном синапсе амплитуда ВПСП значительно меньше и недостаточна для того, чтобы деполяризовать мембрану нейрона до КУД. Поэтому для генерации ПД в нервной клетке требуется возникновение нескольких ВПСП. ВПСП, образовавшиеся в результате срабатывания разных синапсов, электротонически распространяются по мембране клетки, суммируются и генерируют образование ПД в области аксонного холмика. Мембрана нейрона в области аксонного холмика обладает низким электрическим сопротивлением и имеет большое количество потенциалчувствительных Na+ -каналов.
Особенности работы тормозного химического синапса
В тормозном химическом синапсе молекулы медиатора, взаимодействуя с рецепторами постсинаптической мембраны, вызывают открытие К+ — и Cl – -хемочувствительных каналов. Вход в клетку Cl– и дополнительная утечка из клетки К+ приводят к гиперполяризации постсинаптической мембраны, которую называют тормозным постсинаптическим потенциалом (ТПСП) . Возникшая гиперполяризация, во-первых, снижает возбудимость клетки. Во-вторых, ТПСП может нейтрализовать возникший в другом месте клетки ВПСП.
Свойства синапсов
Сравнительная характеристика свойств электрических и химических синапсов приведена в табл. 1.
Одностороннее проведение возбуждения в химическом синапсе связано с его функциональной асимметрией: молекулы медиатора выделяются только на пресинаптической мембране, а рецепторы медиатора расположены только на постсинаптической мембране.
Высокая утомляемость химического синапса объясняется истощением запасов медиатора. Утомляемость электрического синапса соответствует утомляемости нервного волокна.
Низкая лабильность химического синапса определяется главным образом периодом рефрактерности хемочувствительных каналов на постсинаптической мембране.
Синаптическая задержка – время от момента возникновения возбуждения в пресинаптической мембране до момента возникновения возбуждения в постсинаптической мембране. Относительно длительное время синаптической задержки в химическом синапсе (0,2–0,7 мс) затрачивается на вход Са++ в синаптическое окончание, экзоцитоз, диффузию медиатора.
Чувствительность синапса к внешним воздействиям определяется характером процессов, протекающих в синапсе при передаче возбуждения. Химические синапсы чувствительны к действию химических веществ, влияющих на синтез и секрецию медиатора, взаимодействие медиатора с рецептором.
Таблица 1.Свойства электрических и химических синапсов
Источник
Передача информации в химических синапсах
МЕХАНИЗМ СИНАПТИЧЕСКОГО ПРОВЕДЕНИЯ
План лекции:
Понятие синапса. Виды синапсов;
Особенности строения и функционирования химических синапсов;
· передача информации в химических синапсах
· квантовое выделение медиаторов
· генерация потенциала действия
Особенности строения и функционирования электрических синапсов;
Понятие синапса. Виды синапсов
Термин синапс (от греческого sy’napsys — соединение, связь) ввел И. Шеррингтон в 1897 году. В настоящее время синапсами называют специализированные функциональные контакты между возбудимыми клетками (нервными, мышечными, секреторными), служащие для передачи и преобразования нервных импульсов. По характеру контактирующих поверхностей различают: аксо-аксональные, аксо-дендритические, аксо-соматические, нервно-мышечные, нейро-капиллярные синапсы. Электронно-микроскопические исследования выявили, что синапсы имеют три основных элемента: пресинаптическую мембрану, постсинаптическую мембрану и синаптическую щель (рис. 5.1).
Рис. 5.1. Основные элементы синапса.
Передача информации через синапс может осуществляться химическим или электрическим путем. Смешанные синапсы сочетают химические и электрические механизмы передачи. В литературе на основании способа передачи информации принято выделять три группы синапсов — химические, электрические и смешанные.
Строение химических синапсов
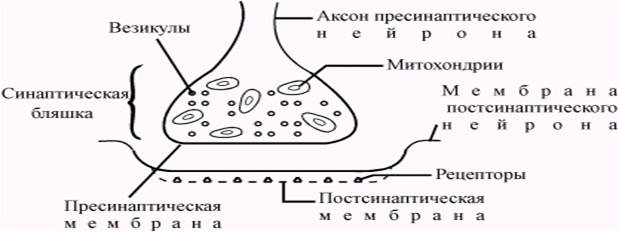
Передача информации в химических синапсах осуществляется через синаптическую щель — область внеклеточного пространства шириной 10-50 нм, разделяющую мембраны пре- и постсинаптических клеток. В пресинаптическом окончании содержатся синаптические везикулы (рис. 5.2) — мембранные пузырьки диаметром около 50 нм., в каждом из которых заключено 1х104 — 5х104 молекул медиатора. Общее количество таких пузырьков в пресинаптических окончаниях составляет несколько тысяч. Цитоплазма синаптической бляшки содержит митохондрии, гладкий эндоплазматический ретикулум, микрофиламенты.
Рис. 5.2. Строение химического синапса
Синаптическая щель заполнена мукополисахаридом, «склеивающим» пре- и постсинаптическую мембраны.
Постсинаптическая мембрана содержит крупные белковые молекулы, выполняющие функции рецепторов, чувствительных к медиатору, а также многочисленные каналы и поры, через которые в постсинаптический нейрон могут поступать ионы.
Передача информации в химических синапсах
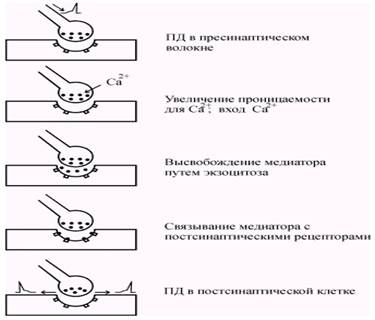
При поступлении потенциала действия к пресинаптическому окончанию происходит деполяризация пресинаптической мембраны и повышается ее проницаемость для ионов Ca 2+ (рис. 5.3). Повышение концентрации ионов Ca 2+ в цитоплазме синаптической бляшки инициирует экзоцитоз везикул наполненных медиатором (рис. 5.4).
Содержимое везикул высвобождается в синаптическую щель, и часть молекул медиатора диффундирует, связываясь с рецепторными молекулами постсинаптической мембраны. В среднем каждая везикула содержит около 3000 молекул медиатора, а диффузия медиатора до постсинаптической мембраны занимает около 0,5 мс.
Рис. 5.3. Последовательность событий, происходящих в химическом синапсе от момента возбуждения пресинаптического окончания до возникновения ПД в постсинаптической мембране.
Рис. 5.4. Экзоцитоз синаптических пузырьков с медиатором. Пузырьки сливаются с плазматической мембраной и выбрасывают свое содержимое в синаптическую щель. Медиатор диффундирует к постсинаптической мембране и связывается с расположенными на ней рецепторами. (Экклз, 1965).
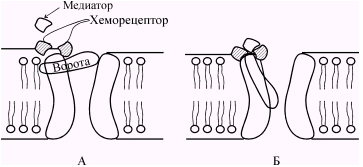
При связывании молекул медиатора с рецептором его конфигурация изменяется, что приводит к открытию ионных каналов (рис. 5.5) и поступлению через постсинаптическую мембрану в клетку ионов, вызывающих развитие потенциала концевой пластинки (ПКП). ПКП есть результат местного изменения проницаемости постсинаптической мембраны для ионов Na + и К + . Но ПКП не активирует другие хемовозбудимые каналы постсинаптической мембраны и его величина зависит от концентрации медиатора, действующего на мембрану: чем больше концентрация медиатора, тем выше (до определенного предела) ПКП. Таким образом, ПКП в отличие от потенциала действия градуален. В этом отношении он схож с локальным ответом, хотя механизм его возникновения иной. При достижении ПКП некоторой пороговой величины возникают местные токи между участком деполяризованной постсинаптической мембраны и соседними с ней участками электровозбудимой мембраны, что вызывает генерацию потенциала действия.
Рис. 5.5. Строение и работа хемовозбудимого ионного канала. Канал образован макромолекулой белка, погруженной в липидный би слой мембраны. До взаимодействия молекулы медиатора с рецептором ворота закрыты (А). Они открываются при связывании медиатора с рецептором (Б). ( По Ходорову Б.И.).
Таким образом, процесс передачи возбуждения через химический синапс может быть схематически представлен в виде следующей цепи явлений: потенциал действия на пресинаптической мембране поступление ионов Ca 2+ внутрь нервного окончания освобождение медиатора диффузия медиатора через синаптическую щель к постсинаптической мембране взаимодействие медиатора с рецептором активация хемовозбудимых каналов постсинаптической мембраны возникновение потенциала концевой пластинки критическая деполяризация постсинаптической электровозбудимой мембраны генерация потенциала действия.
Химические синапсы имеют два общих свойства:
1. Возбуждение через химический синапс передается только в одном направлении — от пресинаптической мембраны к постсинаптической мембране (одностороннее проведение).
2. Возбуждение проводится через синапс значительно медленнее, чем по нервному волокну синаптическая задержка.
Односторонность проведения обусловлена высвобождением медиатора из пресинаптической мембраны и локализацией рецепторов на постсинаптической мембране. Замедление проведения через синапс (синаптическая задержка) возникает вследствие того, что проведение является многоэтапным процессом (секреция медиатора, диффузия медиатора к постсинаптической мембране, активация хеморецепторов, рост ПКП до пороговой величины) и для протекания каждого из перечисленных этапов требуется время. Кроме этого, наличие относительно широкой синаптической щели препятствует проведению импульса с помощью локальных токов.
Химические медиаторы
Медиаторы (от латинского — mediator — проводник) — биологически активные вещества, посредством которых осуществляются межклеточные взаимодействия в синапсах.
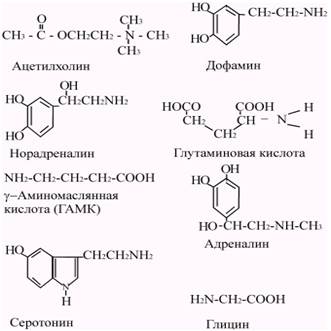
В основном химическими медиаторами являются низкомолекулярные вещества. Однако некоторые высокомолекулярные соединения, такие как полипептиды, могут также выполнить роль химических посредников. В настоящее время известен ряд веществ, играющих роль медиаторов в ЦНС млекопитающих. К ним относятся ацетилхолин, биогенные амины: адреналин, норадреналин, дофамин, серотонин, кислые аминокислоты: глицины, гамма-аминомаслянная кислота (ГАМК), полипептиды: вещество Р, энкефалин, соматостатин и др. (Рис. 5.6).
Рис. 5.6. Структурные формулы некоторых медиаторов.
Функцию медиаторов могут выполнять и такие соединения как АТФ, гистамин, простагландины. В 1935 году Г.Дейлом было сформулированно правило (принцип Дейла), согласно которому каждая нервная клетка выделяет только один определенный медиатор. Поэтому принято обозначать нейроны по типу медиатора, который выделяется в их окончаниях. Так, нейроны, освобождающие ацетилхолин, называется холинергическими, норадреналин — адренергическими, серотонин — серотонинергическими, амины — аминергическими и т.д.





 около 0,5 мс.
около 0,5 мс.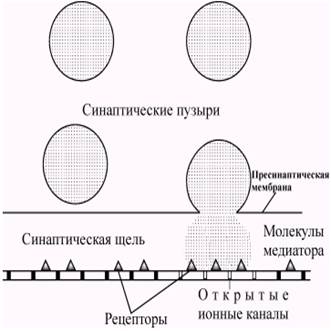
 В основном химическими медиаторами являются низкомолекулярные вещества. Однако некоторые высокомолекулярные соединения, такие как полипептиды, могут также выполнить роль химических посредников. В настоящее время известен ряд веществ, играющих роль медиаторов в ЦНС млекопитающих. К ним относятся ацетилхолин, биогенные амины: адреналин, норадреналин, дофамин, серотонин, кислые аминокислоты: глицины, гамма-аминомаслянная кислота (ГАМК), полипептиды: вещество Р, энкефалин, соматостатин и др. (Рис. 5.6).
В основном химическими медиаторами являются низкомолекулярные вещества. Однако некоторые высокомолекулярные соединения, такие как полипептиды, могут также выполнить роль химических посредников. В настоящее время известен ряд веществ, играющих роль медиаторов в ЦНС млекопитающих. К ним относятся ацетилхолин, биогенные амины: адреналин, норадреналин, дофамин, серотонин, кислые аминокислоты: глицины, гамма-аминомаслянная кислота (ГАМК), полипептиды: вещество Р, энкефалин, соматостатин и др. (Рис. 5.6).


