
База знаний
13.3. Основные пути и способы видообразования
В территориальном аспекте в пространстве новый вид может возникнуть из одной или группы смежных популяций, расположенных на периферии ареала исходного вида. Такое видообразование называется аллопатрическим (от греч. alios — иной, patris — родина). В других случаях новый вид может возникнуть внутри ареала исходного вида, как бы внутри вида; этот путь видообразования называется симпатрическим (от греч. sym — вместе, patris — родина). В филогенетическом аспекте (во времени) новый вид может возникнуть посредством постепенного изменения одного и того же вида во времени, без какой-либо дивергенции исходных групп. Такое видообразование называется филетическим.
Новый вид может возникнуть путем разделения единого предкового вида (дивергентное видообразование). Наконец, новый вид может возникнуть в результате гибридизации двух уже существующих видов — гибридогенное видообразование. Кратко опишем эти основные пути видообразования.
Аллопатрическое видообразование (называемое иногда географическим) иллюстрируется приведенными выше примерами возникновения вида у больших чаек и в группе австралийских мухоловок. При аллопатрическом видообразовании новые виды могут возникнуть путем фрагментации, распадения ареала широко распространенного родительского вида. Примером такого процесса может быть возникновение видов ландыша (см. гл. 6). Другой способ аллопатрического видообразования — видообразование при расселении исходного вида, в процессе которого все более удаленные от центра расселения периферийные популяции и их группы, интенсивно преобразуясь в новых условиях, становятся родоначальниками видов. Примеры, аналогичные видообразованию в группе больших чаек, известны и для других птиц, некоторых рептилий, амфибий, насекомых.
В основе аллопатрического видообразования лежат те или иные формы пространственной изоляции, и этот путь видообразования всегда сравнительно медленный, происходящий на протяжении сотен тысяч поколений. Именно за такие длительные промежутки времени в изолированных частях населения вида вырабатываются те биологические особенности, которые приводят к репродуктивной самостоятельности даже при нарушении первичной изолирующей преграды. Аллопатрическое видообразование всегда связано с историей формирования видового ареала.
Симпатрическое видообразование. При видообразовании симпатрическим путем новый вид возникает внутри ареала исходного вида.
Первый способ симпатрического видообразования — возникновение новых видов при быстром изменении кариотипа, например при автополиплоидии. Известны группы близких видов (обычно растений) с кратными числами хромосом (см. рис. 6.28). Так, например, в роде хризантем (Chrysanthemum) все формы имеют число хромосом, кратное 9, 18, 27, 36, 45, . 90. В родах табака (Nicotiana) и картофеля (Solanum) основное, исходное, число хромосом равно 12, но имеются формы с 24, 48, 72 хромосомами. В таких случаях можно предположить, что видообразование шло путем автополиплоидии — посредством удвоения, утроения, учетверения и т. д. основного набора хромосом предковых видов. Процессы полиплоидизации хорошо воспроизводятся в эксперименте посредством задержки расхождения хромосом в мейозе в результате воздействия, например, колхицином. Известно, что полиплоиды могут возникать и в природных условиях. Возникшие полиплоидные особи могут давать жизнеспособное потомство лишь при скрещивании с особями, несущими то же число хромосом (или при самоопылении). В течение немногих поколений в том случае, если полиплоидные формы успешно проходят «контроль» естественного отбора и оказываются лучше исходной диплоидной, они могут распространиться и сосуществовать совместно с породившим их видом (рис. 13.4) или, что бывает чаще, просто вытеснить его.
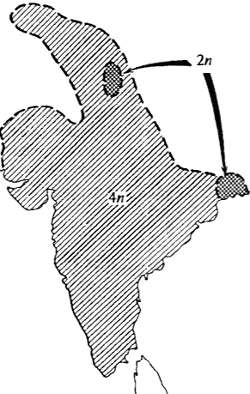
Рис. 13.4. Пример возникновения репродуктивной изоляции при полиплоидизации: обитающий по всему Индостану тетраплоидный вид растений Dicanthium annulatum, несомненно, возник из диплоидной предковой формы, ныне занимающей небольшой дизъюнктный ареал (по Н. Россу, 1962)
Полиплоидные формы, как правило, крупнее и способны существовать в более суровых физико-географических условиях. Именно поэтому в высокогорьях и в Арктике число полиплоидных видов растений резко увеличено (рис. 13.5). Среди животных полиплоидия при видообразовании играет несравненно меньшую роль, чем у растений, и во всех случаях связана с партеногенетическим способом размножения (например, у иглокожих, членистоногих, аннелид и других беспозвоночных).
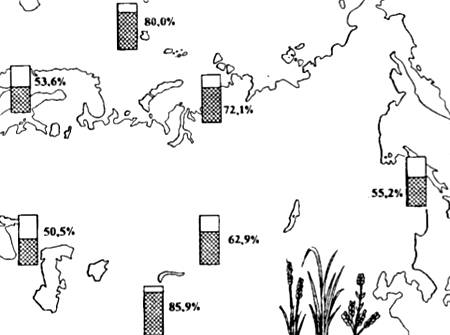
Рис. 13.5. Распространение полиплоидных видов цветковых (в процентах к общему числу видов флоры) в разных частях Евразии (по данным разных авторов из Н.В. Тимофеева-Ресовского и др., 1977)
Второй способ симпатрического видообразования — путем гибридизации с последующим удвоением числа хромосом — аллополиплоидия (см. ниже).
Наконец, последним, достаточно изученным способом симпатрического видообразования является возникновение новых форм в результате сезонной изоляции. Известно существование ярко выраженных сезонных рас у растений, например у погремка Alectorolophus major (см. гл. 10), раннецветущие и позднецветущие формы которого полностью репродуктивно изолированы друг от друга, и, если условие отбора сохраняется, лишь вопрос времени — когда эти формы приобретут ранг новых видов. Аналогично положение с яровыми и озимыми расами проходных рыб; возможно, что эти формы уже являются разными видами, очень схожими морфологически, но изолированными генетически (виды-двойники).
Особенность симпатрического пути видообразования — возникновение новых видов, морфофизиологически близких к исходному виду. Так, при полиплоидии увеличиваются размеры, но общий облик растений сохраняется, как правило, неизменным; при хромосомных перестройках наблюдается та же картина; при экологической (сезонной) изоляции возникающие формы также обычно оказываются морфологически слабо различимыми. Лишь в случае гибридогенного возникновения видов появляется новая видовая форма, отличная от каждого из родительских видов (но также имеющая признаки, характерные для двух исходных видов).
Филетическое видообразование. При филетическом видообразовании вид, весь в целом изменяясь в чреде поколений, превращается в новый вид, который можно выделить, сопоставляя морфологические характеристики этих групп.
Филетическое видообразование включает стазигенез — развитие вида во времени с постепенным изменением одной и той же экологической ниши, и анагенез — развитие вида с приобретением каких-то новых принципиальных приспособлений, позволяющих ему образовать совершенно новую, более широкую экологическую нишу. Примером стазигенеза может быть развитие основного ствола верхнеплиоценовых моллюсков рода Giraulus (см. рис. 6.5).
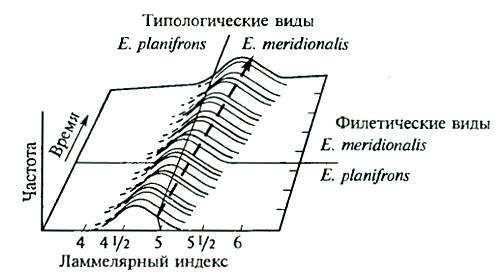
Рис. 13.6. Пример филетического видообразования в ряду ископаемых европейских слонов (Elephas planifrons — Е. meridionalis) по ламеллярному индексу (количеству эмали на зубах) (из В. Гранта, 1980)
Ясно, что в этом случае возможно лишь сопоставление морфологических характеристик, так как изучать результаты филетической эволюции можно лишь с привлечением палеонтологического материала (рис. 13.6). При этом всегда остается возможность того, что на каком-то этапе эволюции от единого филетического ствола могли дивергировать другие группы и филетическое видообразование могло оказаться на самом деле дивергентным. Поэтому в «чистом виде» филетическая эволюция, видимо, возможна лишь как идеализированное и упрощенное отражение эволюционного процесса на одном из отрезков жизни вида (фратрии).
Заметим, что границы между отдельными видами в филетическом ряду форм провести невозможно — она всегда будет условной (см. гл. 12).
Дивергентное видообразование (кладогенез). Ч. Дарвин считал этот тип видообразования самым распространенным (единственный рисунок в «Происхождении видов» посвящен именно этому типу видообразования). Примерами этого типа видообразования — возникновение новых видов в результате разделения единой предковой формы — являются возникновение нескольких видов сигов вокруг Ирландского моря (см. рис. 6.10) и дивергенция дарвиновых вьюрков на Галапагосах (см. рис. 6.12) и эволюция североамериканских дрозофил группы pseudoobscura — persimilis (см. рис. 6.27).
Гибридогенное видообразование (синтезогенез или сингенез). Этот тип видообразования обычен у растений: по некоторым подсчетам, более 50% видов растений представляют собой гибридогенные формы — аллополиплоиды. Укажем лишь на некоторые. Культурная слива (Prunus domestica) с 2n = 48 возникла путем гибридизации терна (P. spinosa, 2n = 32) с алычой (P. divaricata, 2n= 16) с последующим удвоением числа хромосом. Некоторые виды пикульника, малины, табака, брюквы, полыни, ириса и других растений — такие же аллополиплоиды гибридогенного происхождения.
Интересен случай возникновения нового симпатрического вида у Spartina townsendii (2л = 120) на основе гибридизации с последующим удвоением числа хромосом местного английского вида S. stricta (2n = 50) и завезенной в 70-х годах XIX в. из Северной Америки S. alternifolia (2n = 70). Сейчас ареал этого вида интенсивно расширяется за счет сокращения ареала местного европейского вида. Другой пример гибридогенного вида, возникшего симпатрически,— рябинокизильник (Sorbocotaneaster), сочетающий признаки рябины и кизильника и распространенный в середине 50-х годов в лесах южной Якутии по берегам среднего течения р. Алдан (К.М. Завадский). В результате гибридогенного видообразования особенно часто могут образовываться комплексы видов (или так называемых полувидов), связанных между собой гибридизацией,— сингамеоны (В. Грант). В случае таких гибридных комплексов иногда бывает трудно обнаружить четкие границы между отдельными видами, хотя виды как устойчивые генетические системы выделяются вполне определенно.
Все четыре основные формы видообразования во времени схематично показаны на рис. 13.7.
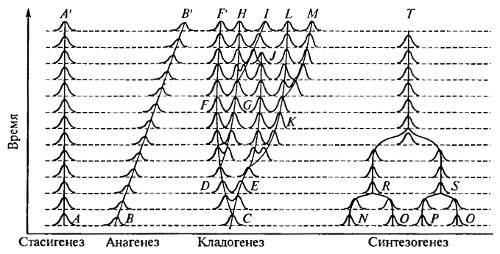
Рис. 13.7. Основные формы филетического (во времени) видообразования (из Н.Н. Воронцова, 2001)
Источник
Гибридогенное видообразование
Наследственность и изменчивость в эволюции.
Биологическая эволюция — необратимое и направленное историческое развитие живой природы, сопровождающееся изменением генетического состава популяций, формированием адаптаций, видообразованием и вымиранием видов, преобразованием экосистем и биосферы в целом.
Наследственность— это свойство организмов повторять в ряду поколений сходные типы обмена веществ и индивидуального развития в целом. Эволюция организмов происходит посредством изменения наследственных признаков организма. Примером наследственного признака у человека может служить коричневый цвет глаз, унаследованный от одного из родителей. Наследственные признаки контролируются генами. Совокупность всех генов организма образует его генотип. Полный набор поведенческих и структурных черт организма называют фенотипом. Фенотип организма образуется за счет взаимодействия генотипа с окружающей средой. Многие черты фенотипа являются ненаследственными. Так, например, загар не наследуется, поскольку его появление обусловлено воздействием солнечного света. Однако, у некоторых людей загар появляется легче, чем у других. Это является наследственной чертой. Передача наследственных признаков от одного поколения к другому обеспечивается ДНК. ДНК — это биополимер, состоящий из четырех нуклеиновых оснований. Во время деления клетки ДНК копируется — в результате каждая из дочерних клеток получает последовательность ДНК.
Части молекулы ДНК, определяющие функциональную единицу наследственности, называются генами. Внутри клеток ДНК находится в составе хроматина, который в свою очередь образует хромосомы. Положение генов на хромосоме называется локусом. Различные формы генов, расположенные в одинаковых локусах гомологичных хромосом и определяющие различные проявления признаков называются аллелями. Последовательность ДНК может изменятся (мутировать), создавая новые аллели. Если мутация происходит внутри гена, то новая аллель может затронуть признак управляемых геном и изменить фенотип организма. Однако большинство признаков определяется не одним геном, взаимодействием нескольких генов (примером подобных явлений могут служить эпистаз и полимерия). Исследование подобных взаимодействий генов одна из главных задач современой генетики. Другим важным вопросом является роль эпигенетических факторов в эволюции.
Изменчивость.Фенотип организма обусловлен его генотипом и влиянием окружающей среды. Существенная часть вариаций фенотипов в популяциях вызвана в различиях их генотипов. Эволюция определяется как изменение с течением времени генетической структуры популяций. Частота одной из аллелей изменяется, становясь более-менее распространенной среди других форм этого гена. Действующие силы эволюции ведут к изменениям в частоте аллели в одну или в другую стороны. Изменение исчезает, когда новая аллель достигает точки фиксации — целиком заменяет предковую аллель или исчезает из популяции.Изменчивость складывается из мутаций, потока генов и рекомбинации генетического материала. Изменчивость также увеличивается за счет обменов генами между разными видами, таких как горизонтальный перенос генов у бактерий, гибридизация у растений. Несмотря на постоянные увеличение изменчивости за счет этих процессов, большая часть генома идентична у всех представителей данного вида. Однако даже сравнительно небольшие изменения в генотипе могут вызвать огромные различия в фенотипе, например, геномы шимпанзе и людей различаются всего на 5 %.
Борьба за существование
Борьба за существование — один из движущих факторов эволюции, наряду с естественным отбором и наследственной изменчивостью, совокупность многообразных и сложных взаимоотношений, существующих между организмами и условиями среды. Также третья глава книги Чарльза Дарвина «Происхождение видов путём естественного отбора» имеет название «Борьба за существование».
Классификация.Дарвин выделил следующие типа борьбы за существование:
Внутривидовая борьба — которая протекает наиболее остро, так как у всех особей вида совпадает экологическая ниша. В ходе внутривидовой борьбы организмы конкурируют за ограниченные ресурсы — пищевые, территориальные, самцы некоторых животных конкурируют между собой за оплодотворение самки, а также другие ресурсы. Для снижения остроты внутривидовой борьбы организмы вырабатывают различные приспособления — разграничение индивидуальных участков, сложные иерархические отношения. У многих видов организмы на разных этапах развития занимают разные экологические ниши, например, личинки жесткокрылых обитают в почве, а стрекоз — в воде, в то время как взрослые особи заселяют наземно-воздушную среду. Внутривидовая борьба приводит к гибели менее приспособленных особей, способствуя таким образом естественному отбору.
Межвидовая борьба — борьба за существование между разными видами. Как правило, межвидовая борьба протекает особенно остро, если у видов сильно перекрываются экологические ниши (часто у представителей одного рода или семейства). В ходе межвидовой борьбы организмы также конкурируют за одни и те же ресурсы — пищевые, территориальные. Межвидовая борьба за существование включает в себя отношения типа хищник — жертва, паразит — хозяин, растение — травоядное животное. Межвидовая борьба за существование во многих случаях стимулирует эволюционные изменения у видов, см. статью Гипотеза Красной королевы. Другим примером борьбы за существование являются взаимно полезное влияние одного вида на другой или другие (например, мутуалистические отношения, комменсализм), подобным образом животные опыляют растения и переносят семена, питаясь нектаром, пыльцой и плодами. Часто межвидовая борьба за существование приводит к появлению приспособлений, как, например, в случае коэволюции цветковых растений и насекомых-опылителей. Обычно межвидовая борьба за существование усиливает и обостряет внутривидовую борьбу. Борьба с неблагоприятными условиями окружающей среды — также усиливает внутривидовую борьбу-состязание, так как, кроме борьбы между особями одного вида, появляется также конкуренция за факторы неживой природы — например, минеральные вещества, свет и другие. Наследственная изменчивость, повышающая приспособленность вида к факторам окружающей среды, приводит к биологическому прогрессу.
Видообразование
Видообразование — процесс возникновения новых видов, процесс изменения старых видов и появления новых в результате накопления новых признаков. При этом генетическая несовместимость новообразованных видов, то есть их неспособность производить плодотворное потомство или вообще потомство, при скрещивании называется межвидовым барьером, или барьером межвидовой совместимости. Существуют разнообразные теории, объясняющие механизмы видообразования, ни одна из которых не считается общепризнанной и полностью доказанной. Одна из причин этого — сложность эмпирической проверки из-за долговременности изучаемого процесса. Согласно синтетической теории эволюции (СТЭ), основой для видообразования является наследственная изменчивость организмов, ведущий фактор — естественный отбор.
В СТЭ выделяют два способа видообразования:
— географическое, или аллопатрическое (Вызывается разделением ареала вида на несколько изолированных частей. При этом на каждую такую часть отбор может действовать по-разному, а эффекты дрейфа генов и мутационного процесса будут явно отличаться. Тогда со временем в изолированных частях будут накапливаться новые генотипы и фенотипы. Особи в разных частях ранее единого ареала могут изменить свою экологическую нишу. При таких исторических процессах степень расхождения групп может достигнуть видового уровня);
— экологическое, или симпатрическое (Связано с расхождением групп особей одного вида и обитающих на одном ареале по экологическим признакам. При этом особи с промежуточными характеристиками оказываются менее приспособленными. Расходящиеся группы формируют новые виды.
Симпатрическое видообразование может протекать несколькими способами. Один из них — возникновение новых видов при быстром изменении кариотипа путём полиплоидизации. Известны группы близких видов, обычно растений, с кратным числом хромосом. Другой способ симпатрического видообразования — гибридизация с последующим удвоением числа хромосом. Сейчас известно немало видов, гибридогенное происхождение и характер генома которых может считаться экспериментально доказанным. Третий способ симпатрического видообразования — возникновение репродуктивной изоляции особей внутри первоначально единой популяции в результате фрагментации или слияния хромосом и других хромосомных перестроек. Этот способ распространён как у растений, так и у животных. Особенностью симпатрического пути видообразования является то, что он приводит к возникновению новых видов, всегда морфологически близких к исходному виду. Лишь в случае гибридогенного возникновения видов появляется новая видовая форма, отличная от каждой из родительских);
«Мгновенное» видообразование на основе полиплоидии
Не предполагает деление ареала на части и формально является симпатрическим. При этом за несколько поколений в результате резких изменений в геноме формируется новый вид. Сальтационно происходит видообразование на основе полиплоидии у растений.
Гибридогенное видообразование
При скрещивании различных видов потомство обычно бывает стерильным. Это связано с тем, что число хромосом у разных видов различно. Несходные хромосомы не могут нормально сходиться в пары в процессе мейоза, и образующиеся половые клетки не получают нормального набора хромосом. Однако, если у такого гибрида происходит геномная мутация, вызывающая удвоение числа хромосом, то мейоз протекает нормально и дает нормальные половые клетки. При этом гибридная форма приобретает способность к размножению и утрачивает возможность скрещивания с родительскими формами. Кроме того, межвидовые гибриды растений могут размножатся вегетативным путем. Существующие в природе естественные ряды гибридных видов растений возникли, вероятно, именно таким путем. Так, известны виды пшеницы с 14, 28 и 42 хромосомами, виды роз с 14, 28, 42 и 56 хромосомами и виды фиалок с числом хромосом, кратным 6 в интервале от 12 до 54. По некоторым данным, гибридогенное происхождение имеют не менее трети всех видов цветковых растений. Гибридогенное происхождение доказано и для некоторых видов животных, в частности, скальных ящериц, земноводных и рыб. Некоторые виды кавказских ящериц, имеющих гибридогенное происхождение, триплоидны и размножаются с помощью партеногенеза.
Наблюдение видообразования
Процесс видообразования, согласно синтетической теории эволюции, очень долог, между тем по сальтационным теориям видообразование происходит очень быстро. Поэтому, собственно, процесс видообразования от начала до конца пока никто не наблюдал. Несмотря на множество экспериментов, представления о механизмах видообразования — это теории со слабой практической базой (исключение составляют эксперименты по ресинтезу видов, сформировавшихся путём полиплоидизации). Есть разные группы в пределах одного вида, в разной степени разошедшиеся по каким-либо причинам. Тем не менее, эти группы всё ещё относятся к одному виду. Есть группы очень близких видов, тем не менее их образование произошло довольно давно по историческим меркам, за пределами наблюдений.
Пример.Яблонная муха (Rhagoletis pomonella) является примером ранней стадии симпатрического видообразования. Первоначально вид обитал в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника. Однако с завозом в Америку яблонь (первое упоминание яблонь в Америке — 1647 год), открылась новая экологическая ниша. В 1864 году личинки Rhagoletis pomonella были обнаружены в яблоках, тем самым зафиксирована яблонная раса этого вида. За полтора века наблюдений расы очень сильно разошлись. Они почти не скрещиваются друг с другом (уровень гибридизации не превышает 4—6 %). Яблоневая раса спаривается почти исключительно на яблонях, а боярышниковая — на боярышнике, что, учитывая разное время созревания плодов, приводит к репродуктивной изоляции. В скором времени возможно выделение из этих рас самостоятельных видов.Кроме того вслед за Rhagoletis pomonella подобное видообразование произошло у наездника Diachasma alloeum, личинки которого паразитируют на личинках мух. После появление новой расы мух появилась и новая раса наездников, предпочитающая паразитировать на яблонных мухах.
Дата добавления: 2018-02-15 ; просмотров: 1964 ; Мы поможем в написании вашей работы!
Источник



