
- Гаструляция у ланцетника тип гаструлы способ
- Ланцетник. Развитие ланцетника.
- Гаструляция у ланцетника
- Сущность и способы гаструляции. Гаструляция у различных представителей хордовых
- Период образования зародышевых листков. Сущность стадии гаструляции. Образование ранней гаструлы. Закладка мезодермы и формирование целома. Гаструляция ланцетника, у амфибий, у пресмыкающихся и птиц, у млекопитающих. Презумптивные карты зародышей.
- Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Гаструляция у ланцетника тип гаструлы способ
Гаструляция — результат активного деления клеток, роста и направленных перемещений (миграций) клеточных потоков с формированием многослойного зародыша, или гаструлы, (возникновением послойно расположенных, отделенных друг от друга отчетливой щелью, зародышевых листков: наружного — эктодермы, среднего — мезодермы, внутреннего — энтодермы). Перемещение клеток происходит в строго определенной области зародыша — в области серого серпа. Последний был описан В. Ру в 1888 г. В оплодотворенном яйце амфибии серый серп выявляется как окрашенная область на стороне, противоположной проникновению спермия. В этом месте, как полагают, локализуются факторы, необходимые для гаструляции.
У разных представителей позвоночных гаструляция совершается несколькими основными способами: путем инвагинации (впячивания), иммиграции (перемещения части клеток внутрь зародыша), эпиболии (обрастания), деламинации (расщепления). Способы гаструляции зависят от типа яйцеклетки. При любом способе гаструляции в качестве ведущих сил выступают неравномерная пролиферация клеток в разных частях зародыша, уровень обменных процессов в клетках, расположенных в разных частях зародыша, активность амебоидных движений клеток, а также индуктивные факторы (белки, нуклеопротеиды, стероиды и др.). В результате гаструляции обособляются основные зачатки органов и тканей.
Следующим периодом эмбриогенеза является гисто- и органогенез — дифференцировка различных тканей и органов организма из материала зародышевых листков и эмбриональных зачатков.
В результате гаструляции возникает многослойный зародыш. Несмотря на различные способы гаструляции после выделения материала зародышевых листков по оси зародыша находится материал хорды, который подстилает нервную пластинку, слева и справа от хорды располагается материал мезодермы. Все это характеризует осевой комплекс зачатков. В дальнейшем происходит формирование зачатков органов, представляющих собой пространственно локализованные группы стволовых клеток — источников развития тканей. Закономерности дифференцировки клеточного материала зачатков можно проследить в эмбриогенезе наиболее изученных животных.
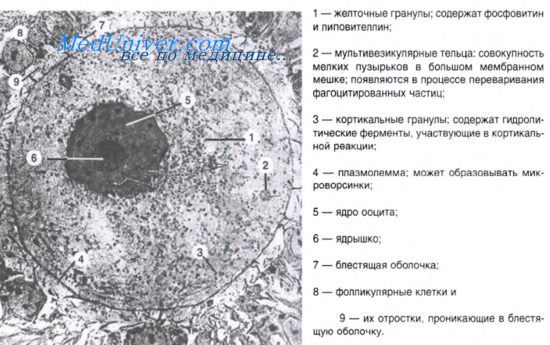
Ланцетник. Развитие ланцетника.
Классическим объектом эмбриологических исследований ланцентник, подробно изученный А.О. Ковалевским. Ланцетник является представителем класса хордовых животных подтипа бесчерепных, размером до 8 см и обитает на песчаном дне в теплых морях. Свое название получил из-за формы, напоминающей ланцет (хирургический инструмент с обоюдоострым лезвием, современный скальпель).
Яйцеклетка ланцетника олиго- и изолецитальная, размером 110 мкм, ядро располагается ближе к анимальному полюсу. Оплодотворение наружное. Дробление зиготы полное, почти равномерное, синхронное и заканчивается образованием бластулы. В результате чередования меридианальных и широтных борозд дробления формируется однослойная бластула с полостью, заполненной жидкостью — бластоцелем. Бластула сохраняет полярность, ее дно представляет собой вегетативную, а крыша — анимальную части; между ними находится краевая зона.
При гаструляции происходит впячивание вегетативной части бластулы в анимальную. Впячивание постепенно углубляется и, наконец, формируется двустенная чаша с широко зияющим отверстием, ведущим в новообразованную полость зародыша. Такой способ гаструляции называется инвагинацией. Так бластула превращается в гаструлу. В ней материал зародыша оказывается дифференцированным на наружный листок — эктодерму, и внутренний — энтодерму. Полость чаши именуется гастроцелем, или полостью первичной кишки, которая сообщается с внешней средой посредством бластопора, что соответствует заднепроходному отверстию. В бластопоре различают дорсальную, вентральную и две латеральных губы. В результате инвагинации центр тяжести зародыша смещается, и зародыш поворачивается бластопором кверху. Постепенно происходит смыкание краев бластопора и удлинение зародыша. Топография клеток в составе губ бластопора определяет развитие разных частей зародыша. При гаструляции из внутреннего листка гаструлы обособляются хорда и мезодерма, которые располагаются между экто- и энтодермой. Завершается гаструляция образованием осевого комплекса зачатков и дальше — обособлением зачатков органов. Хорда индуцирует развитие нервной трубки из материала дорсальной эктодермы. Эта часть эктодермы утолщается, образуется нервная пластинка (нейроэктодерма), которая прогибается по средней линии и превращается в желобок.
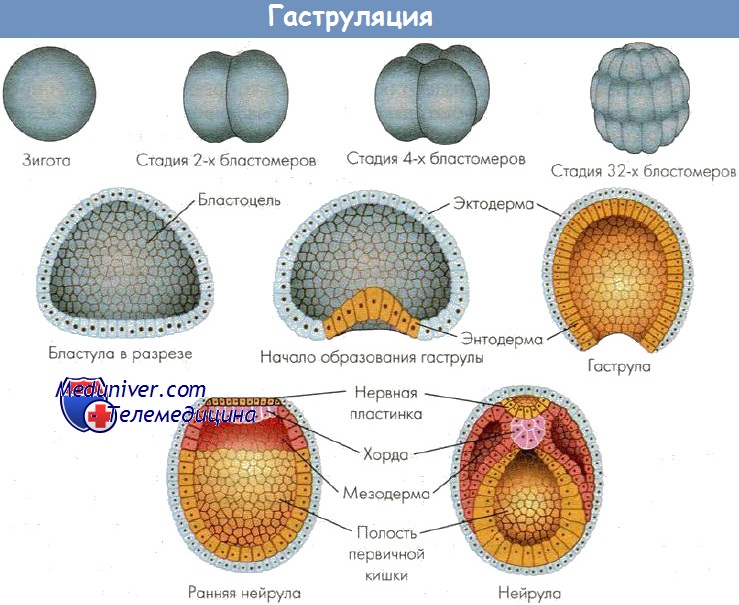
Края желобка постепенно смыкаются в нервную трубку. Оставшаяся часть эктодермы — кожная, срастается над нервной трубкой. Однако на самом переднем и заднем концах зародыша нервная трубка некоторое время сообщается с внешней средой с помощью двух отверстий — нейропоров. В последующем происходит разделение мезодермы на спинные сегменты — сомиты, количество которых нарастает от 15 пар до 60-65 пар у взрослого ланцетника. Часть латерально расположенной мезодермы не сегментируется и расщепляется на наружный (париетальный) и внутренний (висцеральный) листки спланхнотома. Эти листки растут между экто- и энтодермой и, достигнув середины на вентральной стороне зародыша под кишечной трубкой, срастаются, формируя единую вторичную полость — целом. На переднем конце зародыша возникает углубление (ротовая бухта), растущее по направлению к переднему отделу кишечной трубки. При контакте эктодермы ротовой бухты и слепого конца кишечной трубки происходит апоптоз клеток и возникает сообщение кишки с внешней средой. Аналогичный процесс протекает на заднем конце зародыша. По бокам головного отдела зародыша также возникает контакт кожной эктодермы и кишечной энтодермы. В месте этого контакта происходит прорыв. Так полость передней кишки сообщается с внешней средой (формируется жаберный аппарат). После этого зародыш выходит из яйцевой оболочки во внешнюю среду в виде личинки.
Методы маркирования для изучения процессов миграции бластомеров позволили выделить определенные области зародыша на ранних стадиях развития (зиготы — бластулы), которые позднее развиваются в зародышевые листки и эмбриональные зачатки органов и тканей. Эти области были названы презумптивными (предполагаемыми) участками, или зачатками.
Таким образом, на примере развития ланцетника можно наблюдать закономерное перемещение клеток и формирование зародышевых листков, осевого комплекса зачатков и обособление зачатков органов и тканей.
Источник
Гаструляция у ланцетника
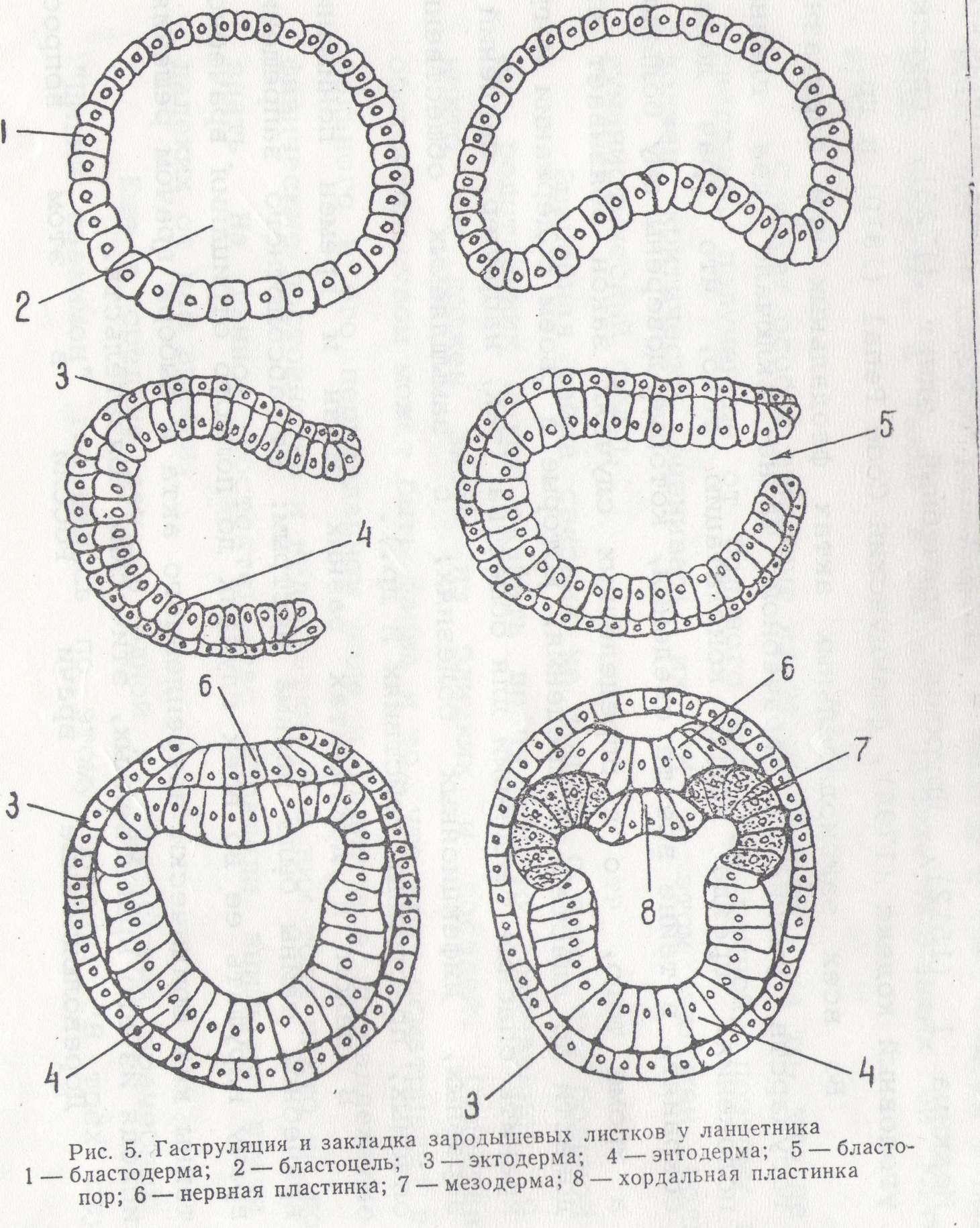
Гаструляция у ланцетника представляет удобную модель для изучения общих закономерностей этого процесса у типа хордовых. Этот процесс, как было показано ранее, у ланцетника, происходит по типу инвагинации. На ранней стадии гаструлы обнаруживается два зародышевых листка: эктодермальный, образованный из крыши бластулы, и энтодермальный— из дна бластулы. Гастроцель сообщается с внешней средой при помощи бластопора.
Наступает следующий этап развития зародыша — образование третьего зародышевого листка—мезодермы. Клетки дорзальной губы бластопора, быстро размножаясь, погружаются в составе энтодермы и на дорзальной ее стороне образуют хордальную пластинку— зачаток будущей хорды. Мелкие клетки вентральной и боковых губ бластопора также врастают в энтодермальный листок и располагаются в его составе по сторонам от хордальной пластинки.
Двуслойный зародыш приобретает вытянутую форму, имея в своем составе материал трех зародышевых листков. Материал мезодермы занимает дорзальную часть внутреннего листка. Из первичной эктодермы, расположенной над хордальной пластинкой, выделяется нервная пластинка. Последняя погружается и по длине зародыша образует нервный желобок. Нервный желобок, замыкаясь кверху, образует нервную трубку.
Хордальная пластинка выходит из состава первичного внутреннего листка, замыкается книзу и образует плотный клеточный тяж — хорду.
Одновременно мелкоклеточный материал, находившийся в первичном внутреннем листке по сторонам от хорды, выпячивается в дорзальном направлении в виде двух карманов. Вначале эти карманы открываются в гастроцель, а затем отделяются в виде двух замкнутых складок, расположенных вдоль гаструлы.
Так образуется третий зародышевый листок — мезодерма. После выделения хорды и мезодермы материал первичного внутренного листка смыкается, образуя энтодерму — внутренний зародышевый листок (рис. 5).
ГАСТРУЛЯЦИЯ у птиц
Гаструляция у птиц представляет довольно близкую модель процессам, имеющим место у высших хордовых, в том числе и у человека.
У птиц в результате дискоидального (меробластического) дробления образуется бластодиск. Он состоит из клеток неправильной формы, плотно прилегающих друг к другу и расположенных на огромной массе нераздробившегося желтка. Под влиянием ферментов клеток бластодиска часть желтка разжижается и образуется полость, заполненная жидкостью — бластоцель. Крыша этой бластулы представлена бластодиском, дно — нераздробившейся массой желтка. Бластоцель — подзародышевая полость.
Затем наступает гаструляция, состоящая из двух фаз. Первая фаза начинается до откладывания яйца и заключается в обособлении энтодермы путем деляминации клеток бластодиска. В результате возникает двуслойный зародыш. Верхний слой клеток дискобластулы приобретает призматическую форму и располагается в один правильный ряд. Нижний слой клеток дискобластулы сохраняет округлую или неправильную форму и лежит беспорядочно на желтке. Между клетками верхнего и нижнего слоя возникает полость — гастроцель.
Вторая фаза гаструляции начинается во второй половине первых суток инкубации. Бластодиск разрастается, в центре его выделяется зародышевый щиток, из которого в дальнейшем развивается тело зародыша. Окружающая зародышевый щиток часть дискобластулы представляет внезародышевый материал, в котором различают светлое и темное поле. Светлое поле расположено вокруг зародышевого щитка и состоит из клеток, отделившихся от желтка подзародышевой полостью в силу частичного использования желтка зародышем. Темное поле занимает периферию дискобластулы и состоит из клеток, плотно прилегающих к желтку и растущих по его поверхности. К этому моменту резко выражено разделение клеточного материала на зародышевую (зародышевый щиток) и внезародышевую (светлое и темное поле) часть.
В течение 24 часов насиживания или инкубации на зародышевом щитке в результате размножения клеток наблюдается перемещение их спереди назад по периферии зародышевого щитка. Оба потока клеток встречаются в центре у заднего конца зародышевого щитка, сливаются и перемещаются кпереди посредине зародышевого щитка. В результате образуется утолщенный тяж клеток, получивший название первичной полоски.
На переднем конце первичной полоски формируется утолщение— первичный узелок. В дальнейшем часть бластомеров, расположенных кпереди от первичного узелка, перемещается к последнему, подворачивается под эктодерму и формирует растущий кпереди между эктодермой и энтодермой головной или хордальный вырост — зачаток хорды.
Бластомеры задней половины зародышевого щитка смещаются к первичной полоске и через эту область погружаются под эктодерму, располагаясь в полости между эктодермой и энтодермой. Этот материал образует зачаток мезодермы — третьего зародышевого листка. Клетки хордального выроста размножаются и. перемещаются, занимая центральное положение между эктодермой
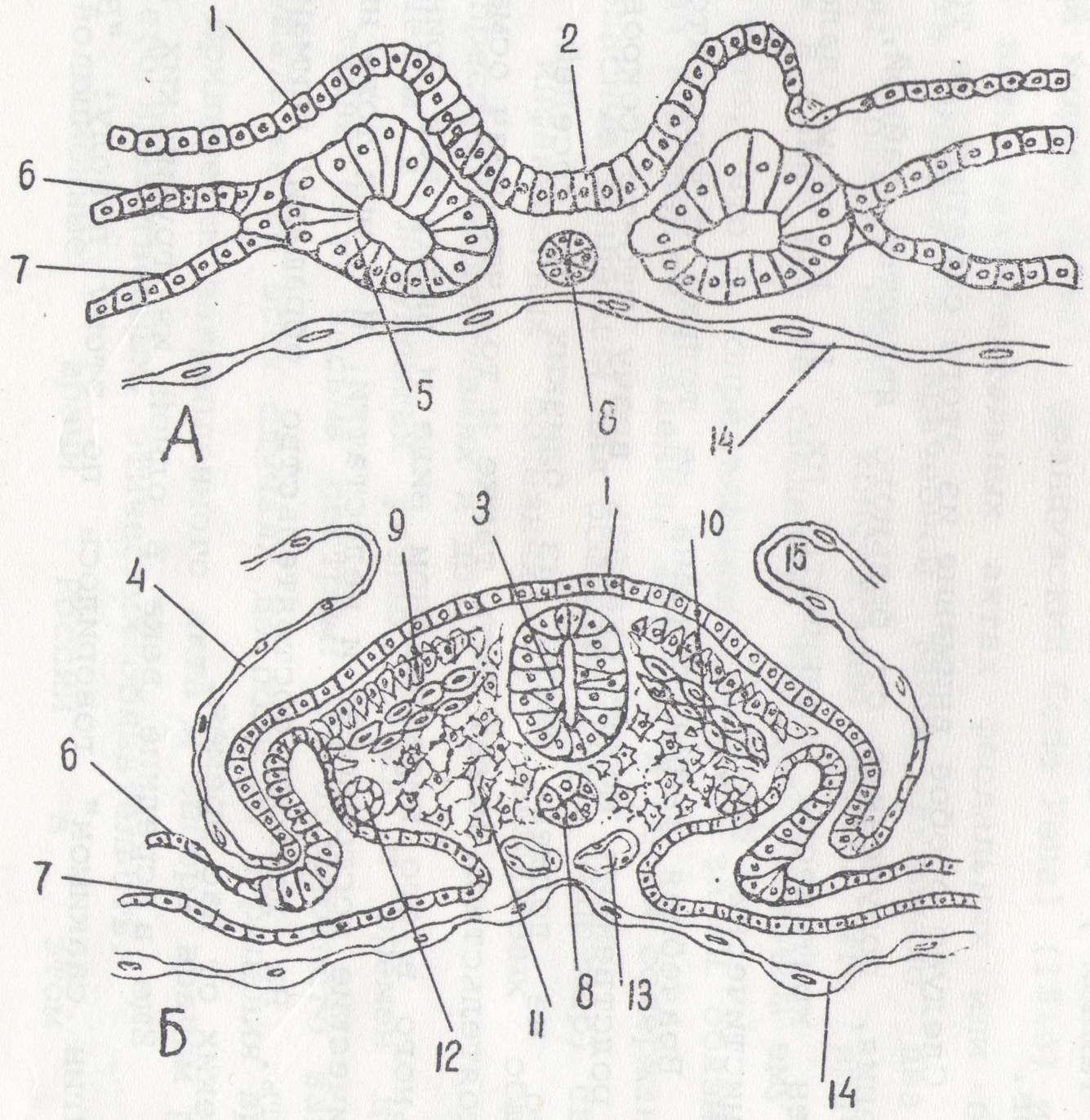
Рис.%6. Развитие комплекса осевых органов у птиц 1—эктодерма; 2 —нервный желобок; 3 — нервная трубка; 4 — амнион; 5 — сомиты; 6 — париетальный листок сплахнотома; 7 — висцеральный листок спланхнотома; 8 — хорда; 9 — дерматом; 10 — миотом; 11 — склеротом; 12 — нефротом; 13 — аорта; 14 — энтодерма; 15 — амниотическая складка и энтодермой в передней половине зародышевого щитка Вслед за этим клетки мезодермы также перемещаются в переднюю половину зародышевого щитка, располагаясь между эктодермой и энтодермой по бокам от хордального выроста. Это парахордальная мезодерма. Ее можно подразделить на дорзальную и вентральную.
По мере того как клетки первичной полоски уходят в закладки хорды и мезодермы на поверхности остается материал эктодермы и входящей в нее нервной пластинки. Таким образом, первичная полоска и головной узелок по своему значению соответствуют бластопору ланцетника, т. к. в этом месте проходит перемещение зародышевого материала, приводящее к образованию 3-зародышевых листков. Наиболее глубоко расположен зачаток кишечной энтодермы. Он непосредственно прилегает к поверхности желтка и находится во внутреннем слое дискобластулы.
К концу гаструляции в теле зародыша имеются все зачатки, из которых возникает типичный для хордовых комплекс осевых органов (рис. 6.).
Источник
Сущность и способы гаструляции. Гаструляция у различных представителей хордовых
Период образования зародышевых листков. Сущность стадии гаструляции. Образование ранней гаструлы. Закладка мезодермы и формирование целома. Гаструляция ланцетника, у амфибий, у пресмыкающихся и птиц, у млекопитающих. Презумптивные карты зародышей.
| Рубрика | Биология и естествознание |
| Вид | курсовая работа |
| Язык | русский |
| Дата добавления | 24.05.2015 |
| Размер файла | 612,8 K |

Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
ФГБОУ ВПО «Пензенский государственный университет»
Педагогический институт им. В.Г. Белинского
Кафедра «Общей биологии и биохимии»
Курсовая работа по дисциплине «Биология»
«Сущность и способы гаструляции. Гаструляция у различных представителей хордовых»
Выполнила: ст. гр. 14ЛС1
1. СПОСОБЫ ГАСТРУЛЯЦИИ
1.1 иммиграция (выселение клеток);
1.2 инвагинация (впячивание);
1.3 эпиболия (обрастание);
1.4 инволюция (вворачивание);
1.5 деляминация (расщепление).
2. ЗАКЛАДКА МЕЗОДЕРМЫ И ФОРМИРОВАНИЕ ЦЕЛОМА
3. ГАСТРУЛЯЦИЯ У РАЗЛИЧНЫХ ПРЕДСТАВИТЕЛЕЙ ХОРДОВЫХ
3.1 Гаструляция ланцетника;
3.2 Гаструляция у амфибий (земноводных);
3.3 Гаструляция у пресмыкающихся и птиц;
3.4 Гаструляция у млекопитающих.
4. ОСОБЕННОСТИ СТАДИИ ГАСТРУЛЯЦИИ
5. ПРЕЗУМПТИВНЫЕ КАРТЫ ЗАРОДЫШЕЙ
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
По окончании периода дробления у многоклеточных животных начинается период образования зародышевых листков — гаструляция.
Сущность стадии гаструляции заключается в том, что однослойный зародыш — бластула — превращается в многослойный — двух- или трехслойный, называемый гаструлой (от греч. «гастер» — желудок в уменьшительном смысле). Гаструляция связана с перемещением эмбрионального материала.
У примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок — эктодерму — и внутренний зародышевый листок — энтодерму. Энтодерма формирует первичную кишку с полостью внутри—гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. Два зародышевых листка являются определяющими морфологическими признаками гаструляции. Их существование на определенной стадии развития у всех многоклеточных животных, начиная с кишечнополостных и кончая высшими позвоночными, позволяет думать о гомологии зародышевых листков и единстве происхождения всех этих животных.
У позвоночных помимо двух упомянутых во время гаструляции образуется еще третий зародышевый листок — мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка, представляющего собой хордомезодерму, является эволюционным усложнением фазы гаструляции у позвоночных и связано с ускорением у них развития на ранних стадиях эмбриогенеза. У более примитивных хордовых животных, таких, как ланцетник, хордомезодерма обычно образуется вначале следующей после гаструляции фазы — органогенезе. Смещение времени развития одних органов относительно других у потомков по сравнению с предковыми группами является проявлением гетерохронии. Изменение времени закладки важнейших органов в процессе эволюции встречается не редко. Процесс гаструляции характеризуется важными, клеточными преобразованиями, такими, как направленные перемещения групп и отдельных клеток, избирательное размножение и сортировка клеток, начало цитодифференцировки и индукционных взаимодействий.
Гаструляция, с которой связаны возникновение различий в клетках и самая ранняя дифференцировка зародыша, сопряжена с экспрессией генов зародыша. Дифференциальную активность генов в процессе гаструляции отражают понятия компетенции и детерминации.
Компетентность — это способность клетки дифференцироваться в нескольких, немногих, направлениях.
Детерминация — это состояние, при котором клетка уже вступила на путь определенной дифференциации и находится в самом его начале.
Согласно ныне существующему определению (Ч. Бодемер, 1971), гаструляция представляет собой ряд морфогенетических движений, в результате которых проспективные зачатки тканей (эктодерма, мезодерма и энтодерма) перемещаются в места, предназначенные для них в соответствии с планом организации [1,стр. 95-101; 3,стр. 104-106].
1. СПОСОБЫ ГАСТРУЛЯЦИИ
Сначала образуется ранняя гаструла, имеющая два зародышевых листка (эктодерму и энтодерму), затем поздняя гаструла, когда формируется третий зародышевый листок — мезодерма. Развитие среднего зародышевого листка можно расценивать как эволюционное усложнение стадии гаструляции у позвоночных, связанное с ускорением у них развития в ранних фазах эмбриогенеза. У примитивных хордовых животных, таких, как ланцетник, аналог 3-го зародышевого листка хордомезодерма обычно образуется в начале следующей после гаструляции стадии — первичного органогенеза. Изменение времени закладки органов и структур в процессе эволюции (гетерохрония) встречается достаточно часто.
Образование ранней гаструлы происходит следующим образом:
1) иммиграцией (выселением клеток), как у кишечнополостных;
2) инвагинацией (впячиванием), как у ланцетника;
3) эпиболией (обрастанием), как у лягушки;
4) инволюцией (вворачиванием);
5) деляминацией (расщеплением), как у некоторых кишечнополостных.
1.1 иммиграция (выселение клеток)
Иммиграции или выселение клеток является наиболее примитивной формой гаструляции. Иммиграция встречается у всех зародышей, но в наибольшей степени характерна для второй фазы гаструляции высших позвоночных.
При этом способе происходит перемещение отдельных клеток или группы клеток из бластодермы в бластоцель с образованием энтодермы, часть клеток бластодермы с поверхности зародыша уходит в бластоцель. Бластоцель заполнена клетками.
Если выселение клеток в бластоцель происходит лишь со стороны одного полюса бластулы, то такая иммиграция называется униполярной, а с различных участков бластулы — мультиполярной. Униполярная иммиграция свойственна некоторым гидроидным полипам, медузам и гидромедузам. В то время как мультиполярная иммиграция является более редким явлением и наблюдается у некоторых гидромедуз. При иммиграции внутренний зародышевый листок — энтодерма может образовываться сразу в процессе проникновения клеток в полость бластоцеля. В других случаях клетки могут заполнять полость сплошной массой, а затем выстраиваться упорядоченно возле эктодермы и образовывать энтодерму. В последнем случае гастроцель появляется позднее (рис. 1).
Рис. 1. Униполярная иммиграция (схема)
1.2 инвагинация (впячивание)
Инвагинация или впячивание наблюдается в случае целобластулы. Это наиболее простой способ гаструляции, при котором вегетативная часть впячивается в бластоцель. Вначале появляется небольшое углубление в вегетативном полюсе бластулы. Затем клетки вегетативного полюса все больше и больше впячиваются в полость бластоцеля. В последующем эти клетки доходят до внутренней стороны анимального полюса. Первичная полость, бластоцель, при этом вытесняется и видна только с двух сторон гаструлы в местах изгиба клеток. Зародыш принимает куполообразную форму и становится двухслойным. Его стенка состоит из наружного листка — эктодермы и внутреннего — энтодермы. В результате гаструляции образуется новая полость — гастроцель или полость первичной кишки. Она сообщается с внешней средой с помощью кольцеобразного отверстия —бластопора или первичного рта. Края бластопора называются губами. Различают спинную, брюшную и две боковых губы бластопора.
По последующей судьбе бластопора всех животных разделяют на две большие группы: первично- и вторичноротых. К первичноротым относят животных, у которых бластопор остается постоянным или дефинитивным ртом у взрослой особи (черви, моллюски, членистоногие). У других животных (иглокожие, хордовые) бластопор или превращается в заднепроходное отверстие, или зарастает, а ротовое отверстие возникает заново на переднем конце тела зародыша. Таких животных называют вторичноротыми. Для ланцетника характерно образование гаструлы путем инвагинации. Впячивание одного из участков бластодермы внутрь осуществляется целым пластом (рис. 2)
Рис. 2. Схема процесса инвагинации:
А — движение пласта клеток; Б — инвагинационная гаструла.
1.3 эпиболия (обрастание)
Эпиболия — это обрастание мелкими быстро делящимися клетками анимального полюса более крупных, отстающих в скорости деления и менее подвижных клеток вегетативного полюса. Такой процесс ярко выражен у земноводных. Эпиболия характерна для животных, развивающихся из телолецитальных яиц. Образование гаструлы идет за счет быстрого деления микромеров, которые обрастают вегетативный полюс. Макромеры оказываются внутри зародыша. Образование бластопора не происходит и нет гастроцели. Эпиболия характерна для амфибий (рис. 3).
Рис. 3. Эпиболия (схема)
1.4 инволюция (вворачивание)
Инволюция — это вворачивание внутрь зародыша увеличивающегося в размерах наружного пласта клеток, который распространяется по внутренней поверхности остающихся снаружи клеток (рис. 4).
Рис. 4. Схема инволюции:
А _ исходный пласт клеток, Б — движения клеточного пласта в ходе инволюции.
1.5 деляминация (расщепление)
Деляминация — расслоение клеток бластодермы на два слоя, лежащих друг над другом. Этот тип гаструляции ограничивается выравниванием внутренних стенок клеток наружного слоя, причем такое выравнивание нередко идет волной от одной соседней клетки к другой. Деляминацию можно наблюдать в дискобластуле зародышей с неполным дроблением, таких, как пресмыкающиеся, птицы, яйцекладущие млекопитающие. Деляминация проявляется в эмбриобласте плацентарных млекопитающих, приводя к образованию гипобласта и эпибласта. Встречается у кишечнополостных. Такой способ гаструляции наблюдается у многих беспозвоночных и высших позвоночных животных (рис. 5).
Рис. 5. Деляминация (схема)
2. закладка мезодермы и формирование целома
Клетки бластодермы делятся на наружный и внутренний слои. Наружный слой образует эктодерму, внутренний — энтодерму. У всех многоклеточных, кроме губок и кишечнополостных, образуется третий зародышевой листок — мезодерма.
Формирование мезодермы происходит двумя способами:
Телобластический способ характерен для первичноротых. На границе между эктодермой и энтодермой по бокам от бластопора клетки телобласты — начинают делиться и дают начало мезодерме. У первичноротых во время гаструляции на границе между эктодермой и энтодермой, по бокам бластопора, уже имеются две большие клетки (или несколько таких клеток) — телобласты, получившие в ходе делений дробления всю полярную ооплазму. Впоследствии они отделяют от себя мелкие клетки и формируют, таким образом, средний пласт — мезодерму. Постепенно телобласты, давая новые и новые поколения клеток мезодермы, отодвигаются к заднему концу зародыша. По этой причине такой способ закладки называют телобластическим.
Энтероцельный способ характерен для вторичноротых. Клетки, формирующие мезодерму, обособляются в виде карманов первичной кишки. Полости карманов превращаются в целом. Мезодерма делится на отдельные участки — сомиты, из которых образуются определенные ткани и органы.
У некоторых животных в связи с увеличением количества желтка в яйцеклетке и уменьшением полости бластоцеля гаструляция только путем инвагинации становится невозможной. Тогда гаструляция происходит способом эпиболии или обрастания. Этот способ состоит в том, что мелкие анимальные клетки усиленно делятся и обрастают вокруг более крупных вегетативных. Мелкие клетки образуют эктодерму, а клетки вегетативного полюса формируют энтодерму. Такой способ гаструляции наблюдается у круглоротых и земноводных [3, стр. 104-106].
При энтероцельном способе мезодерма образуется в виде карманообразных выростов по бокам энтодермы после гаструляции. Эти выпячивания располагаются между экто- и энтодермой, образуя третий зародышевый листок. Такой способ образования мезодермы характерен для иглокожих, ланцетника.
Однако все описанные способы гаструляции редко встречаются отдельно, обычно они комбинируются. Например, вместе с обрастанием может происходить впячивание (земноводные). Расслаивание может наблюдаться вместе с инвагинацией и иммиграцией (рептилии, птицы и др.).
Следовательно, в процессе гаструляции часть клеток из наружного слоя бластулы перемещается внутрь. Это вызвано тем, что в процессе исторического развития одни клетки приспособились к развитию в непосредственной связи с внешней средой, а другие — внутри организма.
3. ГАСТРУЛЯЦИЯ У РАЗЛИЧНЫХ ПРЕДСТАВИТЕЛЕЙ ХОРДОВЫХ
У позвоночных помимо ланцетника во время гаструляции образуется еще третий зародышевый листок — мезодерма, занимающая место между экто- и энтодермой. Развитие среднего зародышевого листка, представляющего собой хордомезодерму, является эволюционным усложнением фазы гаструляции у позвоночных и связано с ускорением у них развития на ранних стадиях эмбриогенеза.
У более примитивных хордовых животных, таких, как ланцетник, хордомезодерма обычно образуется в начале следующей после гаструляции фазы — органогенезе. Смещение времени развития одних органов относительно других у потомков по сравнению с предковыми группами является проявлением гетерохронии.
У представителей разных классов типа Хордовые гаструляция имеет свои особенности. Более детальное рассмотрение гаструляции у ланцетника, земноводных, птиц и млекопитающих поможет глубже понять эволюционные связи и разобраться в закономерностях индивидуального развития.
3.1 Гаструляция у ланцетника
У примитивных хордовых, например у ланцетника, однородная однослойная бластодерма во время гаструляции преобразуется в наружный зародышевый листок —эктодерму —и внутренний зародышевый листок — энтодерму. Энтодерма формирует первичную кишку с полостью внутри—гастроцель. Отверстие, ведущее в гастроцель, называют бластопором или первичным ртом. Два зародышевых листка являются определяющими морфологическими признаками гаструляции. Их существование на определенной стадии развития у всех многоклеточных животных, начиная с кишечнополостных и кончая высшими позвоночными, позволяет думать о гомологии зародышевых листков и единстве происхождения всех этих животных.
У многих беспозвоночных животных (первичноротые) мезодерма образуется из двух клеток — телобластов. Эти клетки обособляются рано, еще на стадии дробления. В процессе гаструляции телобласты располагаются на границе между экто- и энтодермой, начинают активно делиться и образующиеся при этом клетки врастают тяжами между наружным и внутренним листками, образуя мезодерму. Такой способ образования мезодермы называется телобластическим.
Гаструляция у ланцетника происходит путем инвагинации. Разными маркерами на стадии бластулы помечены презумптивные (предполагаемые) зачатки. Это области бластулы, из клеточного материала которых, в ходе гаструляции и раннего органогенеза (нейруляции), обычно образуются совершенно определенные зародышевые листки и органы.
Инвагинация начинается на вегетативном полюсе. Из-за более быстрого деления клетки анимального полюса разрастаются и толкают внутрь бластулы клетки вегетативного полюса. Этому способствует изменение состояния цитоплазмы в клетках, образующих губы бластопора и прилежащих к ним. Вследствие инвагинации бластоцель уменьшается, а гастроцель увеличивается. Одновременно с исчезновением бластоцеля эктодерма и энтодерма приходят в тесный контакт. У ланцетника, как и у всех вторичноротых животных (к ним относят тип Иглокожие, тип Хордовые и некоторые другие относительно малочисленные типы животных), область бластопора превращается в хвостовую часть организма, в отличие от первичноротых, у которых бластопор соответствует головной части. Ротовое отверстие у вторичноротых образуется на противоположном бластопору конце зародыша (рис. 6).
Рис.6 Схема-гаструла ланцетника
1-первичная эктодерма — наружный слой клеток; 2- первичная энтодерма — внутренний слой клеток; 3- бластопор, или первичный рот — щелевидное отверстие; 4- дорсальная губа; 5- вентральная губа; 6-гастроцель, или полость первичного кишечника, — полость, в которую ведёт бластопор; 7-две боковые губы.
3.2 Гаструляция у амфибий (земноводных)
Гаструляция у земноводных имеет много общего с гаструляцией ланцетника, но так как в яйцеклетках у них желтка намного больше и расположен он преимущественно на вегетативном полюсе, крупные бластомеры амфибластулы не способны впячиваться внутрь. Инвагинация проходит несколько иначе, причем настолько, что выделяют особый способ гаструляции (образования экто- и энтодермы) — эпиболию. На границе между анимальным и вегетативным полюсами в области серого серпа клетки сначала сильно вытягиваются внутрь, принимая вид «колбовидных», а затем тянут за собой клетки поверхностного слоя бластулы. Возникают серповидная бороздка и спинная губа бластопора.
У земноводных наблюдается смешанный или переходный способ образования мезодермы. У них мезодерма формируется в процессе гаструляции одновременно с экто- и энтодермой и в ее образовании принимают участие оба зародышевых листка.
Одновременно более мелкие клетки анимального полюса, делящиеся быстрее, начинают перемещаться в сторону вегетативного полюса. В области спинной губы они подворачиваются и впячиваются, а с боков и со стороны, противоположной серповидной бороздке, концентрируются более крупные клетки. По-существу включается процесс эпиболии, который приводит к образованию боковых и брюшной губ бластопора. Бластопор смыкается в кольцо, внутри которого некоторое время видны крупные светлые клетки вегетативного полюса в виде так называемой желточной пробки. Позднее они полностью погружаются внутрь, а бластопор сужается.
На фазу гаструляции приходится начало дифференцировки клеток, что означает переход к активному использованию биологической информации собственного генома. Одним из регуляторов генетической активности является различный химический состав цитоплазмы клеток зародыша, установившийся вследствие ово(оо)плазматической сегрегации. Так, эктодермальные клетки земноводных имеют темный цвет из-за пигмента, попавшего в них из анимального полюса яйцеклетки, а клетки энтодермы — светлый, так как происходят из вегетативного полюса яйца.
С помощью метода маркировки прижизненными (витальными) красителями у земноводных детально изучены перемещения клеток бластулы во время гаструляции, Установлено, что конкретные области бластодермы, называемые презумптивными (лат. рrаеsumрtiо — предположение), при нормальном развитии оказываются сначала в составе определенных зачатков органов, а затем в составе самих органов. Известно, что у бесхвостых амфибий материал презумптивной хорды и мезодермы на стадии бластулы лежит не на ее поверхности, а во внутренних слоях стенки амфибластулы, однако примерно на тех уровнях, как это показано на рисунке (рис. 7).
Определенное сходство процессов гаструляции и областей презумптивных органов у земноводных и ланцетника, т.е. гомология основных органов, таких, как нервная трубка, хорда, вторичная кишка, указывает на их филогенетическое родство.
Рис.7 -Схема — последовательные стадии гаструляции у лягушки.
1 — серповидная бороздка; 2-клетки анимального полюса; 3-вегетативная часть зародыша; 4 — гастроцель; 5 — бластопор; 6 -бластоцель.
3.3 Гаструляция у пресмыкающихся и птиц
У пресмыкающихся, птиц, млекопитающих и человека мезодерма образуется из эктодермы во время второй фазы гаструляции. В течение первой фазы образуется экто- и энтодерма путем деляминации. Во время второй фазы наблюдается иммиграция клеток эктодермы в пространство между экто- и энтодермой. Они и образуют третий зародышевый листок — мезодерму. Этот способ образования мезодермы называется эктодермальным.
Гаструляция у зародышей птиц с меробластическим типом дробления имеет свои особенности. Есть мнение, что она начинается вслед за дроблением и образованием бластулы еще во время прохождения зародыша по яйцеводу. К моменту откладки яйца зародыш как следствие процесса деляминации внутренней клеточной массы бластулы (эмбриобласта в терминологии классической гистологии и эмбриологии) состоит из нескольких слоев: верхний слой называют эпибластом, нижний — первичным гипобластом. Между ними находится узкая щель — бластоцель. Затем образуется вторичный гипобласт, способ образования которого не вполне ясен. Имеются данные о том, что в первичном гипобласте птиц берут начало первичные половые клетки, а вторичный образует внезародышевую энтодерму. Образование первичного и вторичного гипобласта ряд эмбриологов рассматривает как явление, предшествующее гаструляции (во всяком случае основных ее событий).
Основные события гаструляции у птиц начинаются после откладки яиц с началом инкубации. Возникает скопление клеток в задней части эпибласта как результат неравномерного по скорости деления клеток и перемещения их с боковых участков эпибласта к центру, навстречу друг другу. Образуется так называемая первичная полоска, которая вытягивается в направлении головного конца. В центре первичной полоски образуется первичная бороздка, а по краям — первичные валики. На головном конце первичной полоски возникает утолщение — гензеновский узелок, а в нем — первичная ямка [2, стр. 311-316].
Когда клетки эпибласта входят в первичную бороздку, их форма изменяется. Они напоминают по форме «колбовидные» клетки гаструлы земноводных. Затем эти клетки приобретают звездчатую форму и погружаются под эпибласт, образуя мезодерму. Часть мигрирующих клеток, встраиваясь в гипобласт, дает в дальнейшем начало зародышевой энтодерме. Таким образом, энтодерма образуется на основе первичного и вторичного гипобласта с добавлением нового поколения энтодермальных клеток, мигрирующих из верхних слоев бластодермы. Наличие нескольких генераций энтодермальных клеток указывает на растянутость периода гаструляции во времени.
Часть клеток, мигрирующая из эпибласта через гензеновский узелок, образует будущую хорду. Одновременно с закладкой и удлинением хорды гензеновский узелок и первичная полоска постепенно смещаются в направлении от головного к хвостовому концу и исчезают. Это соответствует сужению и закрытию бластопора. По мере сокращения протяженности первичная полоска оставляет за собой сформированные участки осевых органов зародыша в направлении от головных к хвостовым отделам. Представляется обоснованным рассматривать перемещения клеток в курином зародыше как гомологичные таковым у земноводных, а первичную полоску и гензеновский узелок — как структуры, гомологичные спинной (дорзальной) губе бластопора амфибий [9].
Рис. 8- Препарат — гаструляция по типу деляминации и иммиграции. Зародыш птиц (поперечное сечение), стадия первичной полоски. Окраска гематоксилином.
1-первичная полоска; 2-первичная бороздка; 3-эктодерма; 4-мезодерма; 5-энтодерма.
3.4 Гаструляция у млекопитающих
Клетки зародышей млекопитающих, несмотря на то, что у этих животных яйцеклетки имеют малое количество желтка, а дробление полное, в фазе гаструляции сохраняют перемещения, свойственные зародышам пресмыкающихся и птиц. Это подтверждает представление о происхождении млекопитающих от предковой группы, у которой яйца были богаты желтком.
Гаструляция у позвоночных животных, особенно из группы высших, характеризуется разнообразием клеточных процессов. Продолжается, в частности, митотическое размножение клеток, причем оно имеет разную интенсивность в разных частях зародыша. При этом, интерфазы митотических циклов, в отличие от делений дробления, включают все периоды (G1, S, G2), поэтому, начиная со стадии гаструляции, наблюдается рост развивающегося организма. Наиболее характерная черта гаструляции, однако, состоит в перемещении клеточных масс. Это приводит к изменению строения зародыша и превращению его из бластулы в гаструлу: происходит сортировка клеток по их принадлежности к разным зародышевым листкам.
Во время гаструляции очень велика роль эмбриональной индукции — взаимодействия между клеточными комплексами (частями) развивающегося зародыша. Показано, что появление первичной полоски у птиц — результат индукционного взаимодействия между гипобластом и эпибластом. Гипобласту присуща полярность. Изменение положения гипобласта по отношению к эпибласту вызывает изменение ориентации первичной полоски.
Рис.9-Деляминация у млекопитающих
1-гипобласт; 2-зародышевый эпибласт; 3-амниотическая эктодерма; 4-полость амниона.
Рис.10-Иммиграция у млекопитающих
1-первичная полоска; 2- первичный (гензеновский) узелок; 3- клетки, образующие энтодерму и мезодерму; 4-эктодерма; 5-гипобласт.
4. ОСОБЕННОСТИ СТАДИИ ГАСТРУЛЯЦИИ
гаструляция мезодерма ланцетник зародыш
Гаструляция характеризуется разнообразными клеточными процессами. Продолжается митотическое размножение клеток, причем оно имеет разную интенсивность в разных частях зародыша. Вместе с тем наиболее характерная черта гаструляции состоит в перемещении клеточных масс. Это приводит к изменению строения зародыша и превращению его из бластулы в гаструлу. Происходит сортировка клеток по их принадлежности к разным зародышевым листкам, внутри которых они «узнают» друг друга.
На фазу гаструляции приходится начало цитодифференцировки, что означает переход к активному использованию биологической информации собственного генома. Одним из регуляторов генетической активности является различный химический состав цитоплазмы клеток зародыша, установившийся вследствие овоплазматической сегрегации. Так, эктодермальные клетки земноводных имеют темный цвет из-за пигмента, попавшего в них из анимального полюса яйцеклетки, а клетки энтодермы — светлый, так как происходят из вегетативного полюса яйца. Во время гаструляции очень велика роль эмбриональной индукции. Показано, что появление первичной полоски у птиц — результат индукционного взаимодействия между гипобластом и эпибластом. Гипобласту присуща полярность. Изменение положения гипобласта по отношению к эпибласту вызывает изменение ориентации первичной полоски.
Следует заметить, что такие проявления целостности зародыша как детерминация, эмбриональная регуляция и интегрированность присущи ему во время гаструляции в той же мере как и во время дробления [2, стр. 317-319].
5. презумптивные карты зародышей
Для составления карт большое значение сыграла предложенная немецким эмбриологом В. Фогтом (1929) методика маркировки частей зародыша.
При этом на поверхность бластулы наносят метки красками или другими веществами и прослеживая движение меток в ходе гаструляции, отмечают на схеме бластулы или ранней гаструлы судьбу каждой меченой точки (рис.6)
В качестве красящих веществ используют нейтральные краски — нильский голубой, нейтральный красный, метиленовый синий и другие.
Современные методы маркировки эмбриональных тканей позволяют получать карты презумптивных зачатков не только для стадии бластулы, но и для более ранних стадий развития. При этом используют инъекции в отдельные бластомеры флуоресцентных красителей, которые затем выявляются у потомков при рассматривании срезов в люминесцентном микроскопе. Таким способом были составлены карты презумптивных зачатков органов для различных представителей животного мира. [8]
Рис. 6. Маркировка частей зародыша (по Vоgt, 1929):
А — Г — последовательные стадии развития зародыша амфибий.
Цифрами указано положение участков эмбриона, окрашенных разными витальными красителями.
На причины гаструляции единого взгляда не существует. Согласно одним взглядам, гаструляция происходит благодаря неравномерному росту клеток в разных частях зародыша. Румблер (1902) объяснял процесс гаструляции изменением формы клеток внутри и снаружи бластулы. Он считал, что клетки имеют клиновидную форму, внутри бластулы шире, а снаружи уже. Существуют взгляды, что гаструляцию может вызывать резкая интенсивность поглощения воды отдельными клетками. Но наблюдения показывают, что эти различия очень невелики.
Гольтфретер (1943) считал, что анимальный полюс бластулы прикрыт тончайшей пленкой и поэтому клетки связаны в единую массу. Клетки вегетативного полюса не связаны между собой, имеют бутылковидную форму, удлиняются и втягиваются внутрь. В передвижении клеток может играть роль степень слипания и характер межклеточных пространств. Существует также мнение, что клетки могут передвигаться, благодаря их способности к амебоидному движению и фагоцитозу. Образование третьего зародышевого листка в процессе эмбрионального развития животных осуществляется четырьмя способами: телобластическим, энтероцельным, эктодермальным и смешанным.
Гаструляция начинается от 14-го дня развития с формирования первичной полоски на поверхности эпибласта. Сначала она выражена слабо, но на 15-16-е сутки ее хорошо видно в виде утолщения с узкой бороздой посередине. Она локализуется в каудальной части посередине зародышевого диска, тянется в краниокаудальному направлении. Краниальная полоска заканчивается круглым утолщением — первичным узлом (узел Гензена), посередине которого углубление — первичная ямка. Клетки эпибласта мигрируют в направлении первичной полоски, здесь они приобретают пляшкоподибнои формы, отделяются от эпибласта и соскальзывает под него. Первые клетки мигрируют через первичную полоску внутрь, смещают гипобласт и формируют зародышевую эндодерму. Позднее мигрируют клетки, которые занимают позицию мижепибластом и новообразованной эндодермой, формируя мезодерму. Клетки, которые остаются в составе эпибласта, образуют эктодерму. Таким образом, в процессе гаструляции эпибласт дает начало всем трем зародышевым листкам эмбриона. В дальнейшем мезодерма разрастается латерально и краниально в виде мезодермального крыльев.
Клетки, инвагинуют в первичную ямку, движутся в краниальном направлении, формируя нотохорду — зачаток осевого скелета зародыша. Пространство между зародышевыми листками заполняет эмбриональная соединительная ткань — мезенхима. По строению он напоминает сетку, образованную клетками звездчатой формы, которые контактируют своими отростками; межклеточные промежутки заполнены основной межклеточным веществом полужидкой консистенции и тонкими фибриллами. В стенке желточного мешка в конце второй недели развития начинают образовываться кровяные островки и зачатки первичных кровеносных сосудов. В амниотическую ножку из задней части кишечной эндодермы врастает пальцеобразным отросток — аллантоиса. Сосуды желточного мешка прорастают в стенку аллантоиса и ворсинки хориона. Последние омывает материнская кровь. Сформированный в результате описанных процессов алантохорион обеспечивает питание и дыхание плода на раннем этапе его развития [5;6].
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Белоусов Л. В. Основы общей эмбриологии. М.: Изд-во МГУ, 1993. С. 95-118.
2. Васильева В.И. Биология. Под ред. В.Н. Ярыгина. — 5-е изд., испр. и доп. — М.: Изд-во Высш. шк., 2003. С.311-319
3. Чебышов Н.В. Биология (Учебник). М.: Изд-во ВУНМЦ, 2000. С.104-106
4. Ярыгин В.Н. Биология. В 2 кн. Кн. 1: Учеб. для медиц. спец. Вузов. М: Изд-во: Изд-во Высш. шк., 2003. С.309-317
Источник



