
Эндоцитоз осуществляется двумя способами
• Для захвата материала из окружающей среды клетка использует несколько различных механизмов
• В последующих превращениях захваченного материала важную роль играют ферменты деградации и низкое значение pH в эндосомах и лизосомах
Эндоцитоз — процесс, при котором эукариотическая клетка захватывает материал из окружающей среды при образовании везикул на плазматической мембране. Во многом являясь противоположностью экзоцитоза, эндоцитоз обладает несколькими функциями:
• поглощение питательных веществ;
• регуляция экспрессии белков на клеточной поверхности, например рецепторов гормонов и переносчиков глюкозы, что дает возможность клетке контролировать процесс захвата лигандов;
• захват и деградация внеклеточного дебриса;
• восстановление мембраны, встроенной в плазматическую мембрану при секреции.
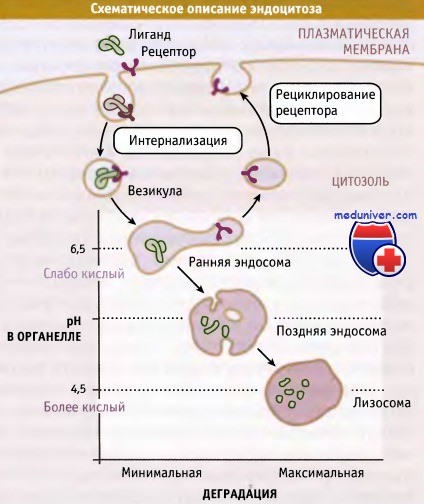 Среди органелл, участвующих в эндоцитозе, существует градиент pH и активности ферментов деградации.
Среди органелл, участвующих в эндоцитозе, существует градиент pH и активности ферментов деградации.
При эндоцитозе макромолекулы или рециклируют в плазматическую мембрану, или разрушаются.
Наряду с этим, процесс эндоцитоза используется для проникновения в клетку патогенными организмами, такими как бактерии, простейшие и вирусы.
При эндоцитозе образующиеся на плазматической мембране везикулы сливаются с органеллами. Эти органеллы обладают двумя важными особенностями: в люмене у них поддерживается кислая среда, и находятся протеолитические ферменты, оптимальная активность которых проявляется при кислых значениях pH.
Органеллы характеризуются различной степенью кислотности среды и различным содержанием ферментов деградации. Они подразделяются на ранние и поздние эндосомы и лизосомы, которые обладают широким спектром протеолитической активности соответственно от минимального в эндосомах до максимального в лизосомах.
В лизосомах находятся ферменты деградации, активные по отношению практически ко всем биополимерам (белкам, липидам, углеводам, РНК, ДНК), попадающим в них путем эндоцитоза. Хотя в течение долгого времени лизосомы рассматривались как терминальные органеллы на пути эндоцитоза, существуют данные о том, что они иногда могут сливаться с плазматической мембраной.
Протонный насос, функционирующий за счет энергии АТФ, называется вакуолярной АТФазой (v-АТФаза) и переносит ионы Н+ из цитозоля в люмен определенных органелл. При этом pH внутри органелл, по сравнению с цитозолем, уменьшается. Значение pH цитозоля составляет 7,4. Внутри ранних эндосом поддерживается слегка кислое значение pH (6,5-6,8), а в поздних эндосомах и в лизосомах значение pH достигает 4,5.
Величина pH в различных эндосомальных компартментах регулируется разными факторами, такими как содержание и активность v-АТФазы, а также ионной проводимостью и свойствами других транспортных АТФаз, присутствующих в мембранах различных эндосом. Важно отметить, что pH в каждом компартменте зависит от выполняемой им функции.
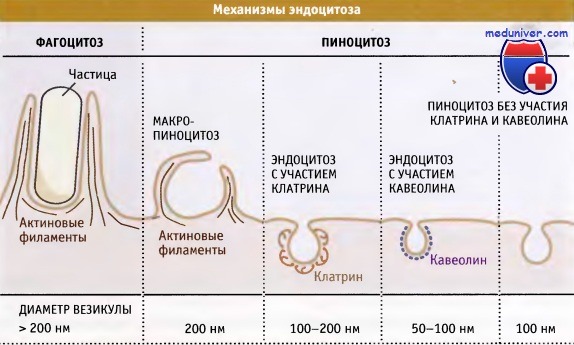 Существует несколько механизмов эндоцитоза.
Существует несколько механизмов эндоцитоза.
Исторически сложилось так, что эндоцитоз стали подразделять на фагоцитоз («поедание клеткой») и пиноцитоз («впитывание клеткой»), что отражает объем попадающего в клетку материала. Специализированный фагоцит, например макрофаг, может заключать материал в везикулу, диаметром до 10 мкм. Сходство с процессом поглощения пищи подчеркивается также сильно кислой внутренней средой фагосом, оптимальной для активности ферментов, расщепляющих поглощенные белки, липиды и углеводы.
Почти все клетки обладают способностью к фагоцитозу, однако этот процесс наиболее характерен для специализированных клеток иммунной системы, таких как макрофаги и дендритные клетки, поглощающих патогенные микроорганизмы и помогающих формировать ся защитной реакции у организма хозяина. Макрофаги также участвуют в очистке организма от стареющих или апоптотических клеток путем фагоцитоза, происходящего даже в отсутствие инфекции. Поскольку настоящие фагоциты обладают специфическими рецепторами, запускающими фагоцитоз, они более эффективны в захвате частиц по сравнению с другими клетками. Например, макрофаги и некоторые другие фагоцитирующие клетки экспрессируют рецепторы для молекул антител.
Пиноцитоз представляет собой общий термин, включающий несколько механизмов захвата. Обычно при этом образуется небольшая эндоцитозная везикула, достигающая 0,1-0,3 мкм в диаметре. Наиболее полно изученной разновидностью пиноцитоза является эндоцитоз, опосредуемый рецепторами. В ранних исследованиях по захвату липопротеинов низкой плотности (ЛНП) соответствующими рецепторами было получено много концептуальных сведений, способствующих нашему пониманию механизмов эндоцитоза. Большое количество рецепторов, расположенных на поверхности клеток, связывает лиганды, к числу которых относятся питательные вещества, ростовые факторы, гормоны, антитела или антигены.
Комплексы «рецептор-лиганд» поглощаются, накапливаясь в специфических областях плазматической мембраны, которые называются «окаймленные ямки». Окаймленные ямки отпочковываются, образуя везикулы, которые сливаются с ранними эндосомами. В кислой среде внутри эндосом связь некоторых лигандов с рецепторами разрывается, причем рецепторы направляются назад, на поверхность клетки, а лиганды — в поздние эндосомы и потом в лизосомы. Некоторые комплексы рецептора и лиганда в ранних эндосомах не диссоциируют, а вместо этого, так же как свободные лиганды, направляются в лизосомы.
Существует несколько вариантов основного пути эндоцитоза. Некоторые клетки содержат лизосомы, в которых происходит лишь частичная деградация поглощенного материала, что существенно для выработки иммунного ответа на захваченные патогены. Хорошим примером являются дендритные клетки, представляющие собой клетки лейкоцитарного происхождения, присутствующие в крови и во всех тканях организма.
Они инициируют почти все иммунные реакции, что обусловлено их уникальной способностью стимулировать В- и Т-лимфоциты, узнающие и уничтожающие патогены. Дендритные клетки захватывают циркулирующие антигены или патогенные микроорганизмы и доставляют их в специальный компартмент лизосом. Эти лизосомы не обладают способностью к полному перевариванию белков, что благоприятствует образованию коротких пептидов (10-15 остатков аминокислот), связывающихся с белками главного комплекса гистосовместимости класса II (МНС). Еще одна особенность этих лизосом позволяет комплексам пептидов и МНС класса II выходить из них за счет образования длинных трубочек, по которым содержимое транспортируется к плазматической мембране. Затем комплексы пептида и МНС класса II выходят на поверхность мембраны, где они стимулируют эффекторные клетки.
 Электронная микрофотография макрофага,
Электронная микрофотография макрофага,
поглощающего эритроцит.
Еще один вариант эндоцитоза предполагает, что поглощенный клеткой материал полностью минует лизосомный путь. Этот процесс носит название трансцитоз, он характерен для специализированных эпителиальных клеток, выстилающих полости тела, например внутреннюю поверхность кишечника.
Функция эпителиальных клеток состоит в регуляции транспорта материалов между организмом и окружающей средой. Эти клетки тесно примыкают друг к другу и образуют непрерывный слой. Они поляризованы, причем «апикальная» поверхность обращена в сторону люмена кишки, а «базальная» — в противоположную сторону, к крови. Обычно трансцитоз начинается с образования покрытой клатрином везикулы на апикальной или базолатеральной мембране. Эта везикула сливается с ранней эндосомой, что приводит к образованию трансцитозной везикулы, принадлежащей к типу рециклирующих везикул. Трансцитозная везикула переносится на противоположную поверхность клетки и сливается с мембраной.
При всасывании в кишечнике питательных веществ некоторые транспортные везикулы, которые образуются при эндоцитозе на апикальной поверхности, продвигаются прямо на базальную сторону и таким образом доставляют интернализированные компоненты без риска их разрушения в лизосомах. Еще одним примером трансцитоза является перенос факторов гуморального иммунитета от матери к новорожденному. Иммуноглобулины, присутствующие в материнском молоке, в кишечнике ребенка захватываются апикальными рецепторами, которые транспортируют их на другую сторону эпителия и доставляют в плазму крови.
Наряду с переносом через окаймленные ямки, на плазматической мембране образуются другие типы эндоцитозных везикул. Кавеолы представляют собой небольшие инвагинации в поверхности мембран, покрытые кавеолином. По-видимому, в кавеолах накапливаются некоторые рецепторы и мембранные липиды, которые не переносятся через окаймленные ямки. Эти рецепторы и липиды попадают в клетку, когда кавеола отпочковывается, образуя везикулу. Плазматическая мембрана также образует более крупные гетерогенные структуры, которые называются макропиносомы.
Обычно они образуются в ответ на действие факторов роста и представляют собой большие вакуоли, похожие по размеру на фагосомы. В них могут содержаться крупные капли внеклеточной жидкости. Материал, попавший в клетку с помощью кавеол и макропиносом, достигает тех же эндосом и лизосом, так же как и материал, поглощенный через окаймленные ямки. В некоторых случаях содержимое кавеол может храниться в специальной эндосоме, которая называется кавеосома. Наконец, некоторые метаболиты попадают в клетку вместе с мелкими пиноцитозными везикулами, которые образуются в отсутствие известных типов оболочек (окаймления).
Интересно, что некоторые вирусы животных используют кислые значения pH, характерные для эндоцитоза, для проникновения в цитоплазму клетки, где они проходят цикл репликации. Так, вирусы везикулярного стоматита и леса Семлики проникают в клетку через кислое окружение люмена эндосом; при этом активируются гликопротеины «шипов» на поверхности вирусной частицы и запускается процесс слияния мембран вируса и эндосомы. При этом происходит попадание в цитоплазму клетки вирусного генома и начинается распространение инфекции.
В клетках почкующихся дрожжей S. cerevisiae происходит пиноцитоз, но не фагоцитоз. Это не вызывает удивления, поскольку дрожжевая клетка обладает толстой клеточной стенкой. У этих клеток плохо различимы органеллы, принимающие участие в эндоцитозе, поскольку их очень мало. Однако вакуоли, которые представляют собой аналоги лизосом, хорошо видны в электронном микроскопе. До недавних пор генетические исследования эндоцитоза на дрожжах отставали от исследований экзоцитоза, и лишь в последнее время они стали особенно важны для понимания роли убиквитинирования белков в процессе их деградации.
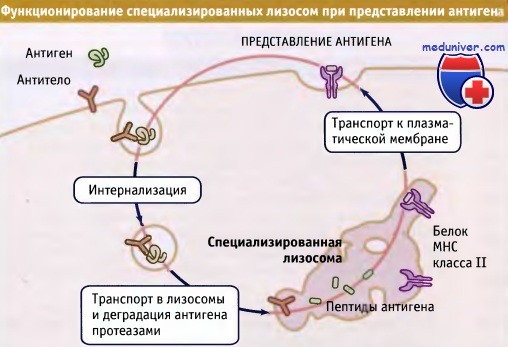 В клетках иммунной системы специализированные лизосомы разрушают захваченные белковые антигены,
В клетках иммунной системы специализированные лизосомы разрушают захваченные белковые антигены,
образуя пептиды, которые связываются с молекулами МНС класса II.
Комплексы пептид-МНС класса II транспортируются в плазматическую мембрану для представления антигена эффекторным клеткам. ![]() В поляризованной клетке процесс трансцитоза протекает таким образом,
В поляризованной клетке процесс трансцитоза протекает таким образом,
что материал, транспортируемый по механизму эндоцитоза из одного мембранного домена,
проходит через клетку и путем экзоцитоза выходит через другой мембранный домен.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в содержание раздела «генетика» на нашем сайте
Источник
ЭНДОЦИТО́З
В книжной версии
Том 35. Москва, 2017, стр. 386
Скопировать библиографическую ссылку:





ЭНДОЦИТО́З (от эндо… , греч. ϰύτος – сосуд, здесь – клетка, и . оз ), процесс захвата и поглощения клеткой каких-либо объектов, которые не способны проникать через плазматич. мембрану. Э. происходит в отношении макромолекул, твёрдых частиц, вирусов, целых клеток и их частей ( фагоцитоз ) или жидкости с растворёнными в ней веществами ( пиноцитоз ). Транспорт в осн. осуществляется в составе везикул (образуются из впячивания на поверхности плазматич. мембраны). При отшнуровывании в цитоплазму они перемещаются к внутр. структурам клетки (напр., лизосомам). С помощью Э. осуществляются питание клетки (для простейших и низших позвоночных это единственный механизм питания), защитная функция (в т. ч. иммунологическая), опосредованная рецепторами регуляция внутриклеточных систем гормонами и др. химич. мессенджерами и т. д. Ср. Экзоцитоз .
Источник
5.7. Экзоцитоз и эндоцитоз
5.7. Экзоцитоз и эндоцитоз
Для клеток различных типов характерны процессы экзо- и эндоцитоза. В приближенном виде экзоцитоз — выделение каких-либо продуктов через плазматическую мембрану во внеклеточную среду, эндоцитоз — поступление материалов из внешней среды в клетку. Эти механизмы служат примером сложных систем, сконструированных из функционально взаимосвязанных блоков, реализующих многоступенчатые процессы.
Экзоцитоз — наиболее распространенный механизм секреции — лежит в основе таких различных феноменов, как внешняя секреция слизи, ферментов, гормонов, выделение нейротрансмиттеров при передаче сигналов от одной клетки к другой и т.д. Этот механизм обеспечивает также выделение альбумина и некоторых других белков гепатоцитами, гистамина — тучными клетками и т.д. При экзоцитозе материал, упакованный в секреторных гранулах, покрытых липопротеиновой мембраной, выбрасывается во внешнюю среду клетки (в ряде случаев в межклеточные пространства) .
Полипептидные компоненты секретируемых материалов синтезируются на рибосомах эндоплазматического ретикулума, затем транспортируются в цистерны аппарата Гольджи, где происходит их упаковка в специальные везикулы. Гранулы секрета формируются преимущественно в результате отшнуровывания кусочков аппарата Гольджи. Предполагается, что по ходу движения к плазматической мембране они сливаются и наполняются секретом. Секреторные гранулы приближаются к внутренней поверхности плазматической мембраны и сливаются с ней. В одной из точек мембраны возникает все увеличивающееся отверстие, через которое выбрасывается содержимое гранул. Далее, мембрана секреторной гранулы превращается в часть плазматической мембраны или вновь погружается внутрь клетки, где образуется пустая везикула, наполняющаяся секретом. Так как при экзоцитозе путь секреторной гранулы не идеален, то та часть секрета, которая поступает через латеральную мембрану, представляет собой продукты внутренней секреции. Напротив, при внутренней секреции незначительная часть секрета выделяется через апикальную мембрану и, следовательно, представляет собой остаточную внешнюю секрецию эндокринных клеток. В пользу этой точки зрения свидетельствует то, что при внешней секреции пищеварительных ферментов клетками поджелудочной железы, желудка, слюнных желез часть этих ферментов поступает в кровь. При патологии соотношение между внешней и внутренней секрецией может меняться.
Эндоцитоз сводится к тому, что под влиянием чаще всего определенного сигнала участок мембраны начинает инвагинироваться внутрь цитоплазмы. Образовавшаяся везикула, покрытая плазматической мембраной, начинает двигаться по направлению к центральным структурам клетки. Можно выделить несколько типов эндоцитозов: 1) фагоцитоз — поступление во внутриклеточную среду «твердых» структур; 2) пиноцитоз — поступление во внутриклеточную среду жидких субстратов; 3) микропиноцитоз, отличающийся от фаго- и пиноцитоза незначительной величиной образовавшихся везикул.
Ранее предполагалось, что все типы эндоцитоза связаны с внутриклеточным пищеварением. Однако в настоящее время ясно, что эндоцитоз играет важную роль в механизмах внутриклеточного транспорта. При этом часть молекул доставляется внутрь клеток как из внешней, так и из внутренней среды. Механизм эндоцитоза не обладает способностью обеспечить нутритивные потребности не только многоклеточных организмов, но и отдельных клеток. Поэтому во многих случаях эндоцитоз выполняет скорее транспортную, чем нутритивную функцию. Эндоцитоз нередко связан с выполнением защитных функций, как это предполагалось в отношении фагоцитоза И. И. Мечниковым. Во многих случаях поглощение бактерий лимфоцитами сопровождается их умерщвлением без глубокого переваривания. Другими словами, защитный фагоцитоз не всегда и не в полной мере представляет собой разновидность внутриклеточного пищеварения.
Особый интерес представляет процесс, происходящий при сочетании эндо- и экзоцитоза. В этом случае образовавшаяся на поверхности мембраны везикула поступает в клетку путем эндоцитоза, а на другом полюсе клетки путем экзоцитоза освобождает свое содержимое. Этот процесс, который мы предлагаем называть трансцитозом, обеспечивает прохождение через клетку частично или полностью пнтактного материала, а также высокомолекулярных соединений. Например, за счет трансцитоза происходит проникновение белков молока матери через кишечный барьер во внутреннюю среду организма ребенка. Вероятно, таким путем обеспечивается иммунная устойчивость организма в течение самых первых дней постэмбрионального периода. Не исключено, что трансцитоз характерен не только для новорожденных, но и для взрослых.
Опосредованный рецепторами эндоцитоз — специализированный механизм (рис. 33), участвующий по крайней мере в 13 различпых процессах интернализации молекул, главным образом белков, в клетках различных типов в норме и при патологии. Рецепторная функция эндоцитоза сводится к тому, что в результате переноса внутрь клетки везикул, образующихся на поверхности клетки, в частности к аппарату Гольджи и другим внутриклеточным органеллам, реализуется транспорт сигналов к внутриклеточным системам. Это особенно важно, когда мембрана клеток непроницаема для этих сигналов. Существуют аргументы, что таким образом передается влияние различных кортикостероидов и других кортикоидных гормонов, инсулина и ряда других. Во всех случаях имеет место проникновение во внутриклеточную среду организма различных белков и пептидов, а иногда и надмолекулярных агрегаций, не проникающих через мембрану с помощью других механизмов.
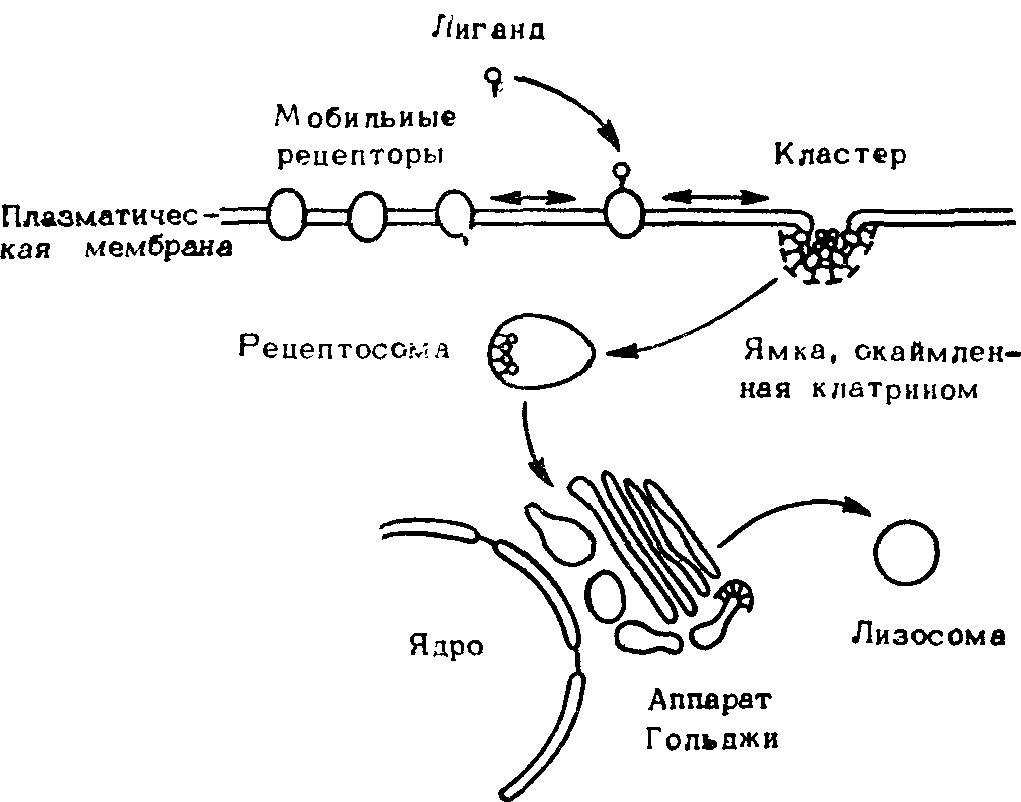
Рис. 33. Упрощенная схема рецептормедиированного эндоцитоза (стадии связывания, накопления и поступления типичных лигандов в фибробласт).
Различные типы рецепторов, синтезированных с участием рибосомного аппарата и во многих случаях подвергающихся последующим процессингам в аппарате Гольджи, встраиваются в плазматическую мембрану с помощью недостаточно ясного механизма. Первоначально их распределение диффузно. При поступлении сигналов — гормонов, нейротрансмиттеров, кейлонов и др. — соответствующие рецепторы образуют комплекс лиганд—рецептор, после чего их способность к латеральному движению возрастает и они начинают концентрироваться в определенных участках клетки—«окаймленных ямках». После того как комплексы лиганд—рецептор в окаймленной ямке достигают определенной концентрации, эта ямка начинает углубляться в цитоплазму, образуя везикулу, которая по механизму возникновения и другим характеристикам отличается от эндоцитозной везикулы. Везикула продвигается внутрь цитоплазмы, на некоторое время соединяется с лизосомами или цистернами аппарата Гольджи и затем вновь возвращается к поверхности мембраны. По-видимому, везикула совершает много оборотов.
При соединении везикулы с лизосомами предполагается разрушение сигнальных молекул или других лигандов при сохранении рецепторов. В случае присоединения к другим органеллам клетки, вероятно, происходит передача сигнала на внутренние структуры клетки. Таким образом, этот процесс, получивший название «интернализация», является еще одним механизмом, с помощью которого реализуется управление клеточными функциями. Выпадение или нарушение эндоцитозного блока будет неизбежно сказываться на всех функциях клетки хотя бы уже из-за нарушений системы сигнализации.
Концепция функциональных блоков встречает многочисленные трудности, но существуют теоретические и эксперименальные аргументы, позволяющие преодолеть их. К одному из затруднений относится вопрос, не противоречит ли концепция универсальных функциональных блоков представлениям об эволюции на молекулярном или близком к нему уровнях. Однако возможность рекомбинации функциональных блоков можно справедливо рассматривать как эволюционный процесс, поразительный по богатству своих возможностей. Примером может служить эволюция генетического аппарата и других информационных систем, где рекомбинация становится основным механизмом. В ряде случаев идея о существовании универсальных функциональных блоков не только не отвергает изменений, но и постулирует их ускоренную эволюцию. Эта же идея позволяет понять случаи консерватизма некоторых молекул или их частей на протяжении огромных отрезков времени.
Другая трудность касается участия значительного числа различных функциональных блоков — изоферментов, изогормонов — в одном и том же механизме. Такое разнообразие блоков также находит объяснение в рамках основной идеи с учетом современных представлений генетики о свойствах генетического аппарата, а цитофизиологии — о посттрансляционных процессингах.
Под давлением фактов следует отказаться от мысли, что специализация и совершенство сложных физиологических функций связаны со специализацией элементарных функций и осуществляющих их функциональных блоков. Напротив, высокая специализация сложных функций достигается на основе рекомбинации и транспозиции большого, хотя и ограниченного набора молекулярных машин, производящих элементарные физиологические операции. Живые системы представляют собой уникальные наборы определенным образом организованных универсальных функциональных блоков. Уникальность и специализация достигаются благодаря сочетаниям универсальных функциональных блоков в пространстве и времени, а не за счет уникальности элементов, из которых строится данная система.
Концепция универсальных функциональных блоков не только влияет на общие представления биологов и врачей, но и приводит к ряду далеко идущих практических заключений. Прежде всего она открывает перспективы для различных способов химического и фармакологического воздействий на организм человека, а также для более серьезных попыток человечества вмешиваться в окружающий его мир для управления им или, напротив, для извлечения определенной пользы, не влияя на природу в целом.
Источник



