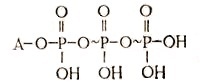Для жизнедеятельности дрожжевой клетки необходим постоянный приток энергии, которую она получает в результате метаболизма (обмена веществ), т. е. превращения веществ.
Процесс питания, или ассимиляция, заключается в поступлении и усвоении пищи. Поскольку вещества пищи отличаются от веществ клетки по химическому составу, они подвергаются сложной переработке. Сначала они расщепляются на макромолекулы: белки, полисахариды, полинуклеотиды, которые в свою очередь распадаются на мономеры — аминокислоты, сахара, жирные кислоты. Эта сторона обмена веществ называется катаболизмом и осуществляется вне клетки под действием экзоферментов.
Образовавшиеся мономеры поглощаются клеткой и из них строятся вещества самой клетки. Этот обмен называется строительным или анаболизмом .
Для процессов метаболизма нужна энергия, которую клетка получает в результате окисления органических веществ. Эта сторона деятельности клетки называется энергетическим обменом , а окисление органических веществ — дыханием (диссимиляция).
Процессы питания и дыхания осуществляются одновременно и составляют две стороны обмена веществ. Энергия, освобождающаяся при дыхании, не выделяется в виде тепла, а аккумулируется в соединениях, богатых энергией, — макроэргических связях АТФ (аденозинтрифосфата).
Углерод . По типу усвоения углерода микроорганизмы делятся на автотрофы и гетеротрофы.
Автотрофы способны синтезировать органические вещества из неорганических. Они делятся на фототрофы и хемотрофы. Фототрофы питаются как зеленые растения, используя для синтеза органических веществ солнечную энергию. Таким образом могут питаться только окрашенные микроорганизмы — синезеленые водоросли и пурпурные серобактерии.
Хемотрофы используют для синтеза органических веществ химическую энергию, освобождающуюся в результате окисления неорганических веществ. Например, нитрифицирующие бактерии окисляют аммиак в нитратную (азотную) кислоту, железобактерии окисляют закисное железо в окисное, бесцветные серобактерии окисляют сероводород до сульфатной (серной) кислоты.
Гетеротрофы питаются готовыми органическими веществами. Они делятся на сапрофиты (органотрофы) и паразиты (паратрофы). Сапрофиты питаются мертвой органической пищей, а паразиты способны питаться веществами живой клетки-хозяина.
Дрожжи по типу питания относятся к сапрофитам. В качестве источника углерода они используют углеводы: дисахариды (сахарозу и мальтозу) и моносахариды — гексозы (глюкозу и фруктозу). Дрожжи не могут усваивать крахмал, клетчатку, пентозы. Лактоза усваивается только дрожжами Saccharomyces lactis (сахаромицес лактис).
Опыты с мечеными атомами углерода показали, что некоторые дрожжи (Saccharomyces cerevisiae — сахаромицес церевизие) способны к ассимиляции углекислого газа как образованного ими при брожении, так и содержащегося в воздухе. Первым продуктом ассимиляции является щавелевоуксусная кислота.
Азот . Дрожжи хорошо усваивают азот как органических, так и неорганических веществ. Из неорганических форм азота они усваивают аммонийные соли фосфорной и сульфатной кислот, а также свободный аммиак. Дрожжи плохо используют нитриты и нитраты. Из органических форм они усваивают аминокислоты, пептиды и мочевину. Усвоение аминокислот сопровождается их дезаминированием и декарбоксилированием. В кислой среде усиливается процесс декарбоксилирования, а в щелочной — дезаминирования.
Дезаминирование может идти двумя путями в зависимости от условий. При окислительном дезаминировании (в присутствии кислорода) образуются кетокислота и аммиак:
R – СН(NH2) – СООН + 0,5О2 → R – СО – СООН + NН3 ↑ .
При восстановительном дезаминировании (в отсутствие кислорода) образуются альдокислота и аммиак:
R – СН(NH2) – СООН + 2Н → R – СН2 – СООН + NН3 ↑ .
При гидролитическом дезаминировании образуются оксикислота и аммиак:
R – СН(NH2) – СООН + Н2О → R – СНОН – СООН + NН3 ↑ .
Опытным путем доказано, что две аминокислоты лучше стимулируют размножение дрожжей и усиливают брожение, чем одна. Из этого напрашивается вывод, что одна аминокислота служит донатором водорода, а другая акцептором:
Смесь аминокислот ускоряет прирост дрожжей: двух аминокислот — на 20 %, трех — на 28, восьми — на 50 % .
Если дрожжи обеспечить всеми необходимыми аминокислотами, то они усваивают их без предварительного дезаминирования. Если в среде отсутствует часть необходимых аминокислот, дрожжи дезаминируют их, чтобы получить аммиак, необходимый для их синтеза.
Фосфор и сера . Фосфор является необходимым элементом для питания дрожжей. Он входит в состав ядра, протоплазмы, митохондрий, рибосом, волютина, содержится в нуклеиновых кислотах, ферментах. Фосфор входит в состав аденозинтрифосфата (АТФ):
обозначены макроэргические связи; при их гидролитическом расщеплении выделяется большое количество энергии (фосфорилирование).
Фосфорилирование — единственный биологический процесс, при помощи которого организмы получают необходимую энергию.
Дрожжи быстрее усваивают неорганический фосфор, чем органический.
Сера необходима дрожжам, как и всем другим живым организмам. Она входит в состав некоторых белков, ферментов в виде сульфгидрильной группы –SH.
Сера нужна дрожжам для синтеза некоторых витаминов (В1) и ростовых веществ (биотина, метионина).
В качестве источника серы дрожжи могут использовать сульфаты, сульфиты, сероводород и элементарную серу. Виноградные и плодовые соки содержат сульфаты. Кроме того, при брожении в соответствии с технологией используется диоксид серы. Может попасть в сусло и элементарная сера, стекающая с серных фитилей при окуривании тары. Элементарную серу дрожжи способны откладывать в своих клетках, но это приводит к изменению их формы.
Источник
Дрожжевые грибы по способу питания являются
Питание дрожжевой клетки.
Питательные вещества либо входят в состав клетки, либо снабжают ее необходимой для жизни энергией.
Перенос питательных веществ через цитоплазматическую мембрану, которая обладает способностью регулировать проникновение различных веществ в клетку и выход из нее, осуществляется в результате переноса двух типов: диффузии и стерео-химического специфичного переноса. Каждый тип переноса имеет активную и пассивную формы (рис. 25).
Процесс пассивной диффузии происходит без затрат энергии клеткой, и вещества проникают через цитоплазматическую мембрану при растворении в ней, а процесс активной диффузии
с затратой энергии (обычно АТФ) в процессе дыхания. Так, для проникновения вещества R—О в клетку потребуется затрата энергии на восстановление его водородом до R — ОН, растворимого в ци-топлааматической мембране, с последующим окислением до R—О в клетке и освобождением водорода для восстановления новой молекулы R—О.
Перенос же в клетку большинства веществ, нерастворимых в цитоплазматической мембране, осуществляется находящимися на мембранах особыми белка-ми-переносчиками — пермеазами. Таким образом, проницаемость цитоплазматической мембраны связана с наличием веществ, роль которых заключается в транспорте ряда веществ в клетку микроорганизма. При пассивном стереохимическом специфичном переносе питательных веществ комплекс вещество — пермеаза растворяется в цитоплазматической мембране, диффундирует в клетку и пермеаза возвращается за новым веществом.
Активный стереохимйческий специфичный перенос питательных веществ требует затраты энергии клеткой микроорганизма на превращение вещества в форму, способную соединиться с белком-носителем и пройти через мембрану. Например, вещество R2—О должно быть превращено в R2—ОН, которое и объединяется со специфической пермеазой [112].
Следует отметить, что эффект переноса растворенных веществ обеспечивает определенная стереохимическая структура пермеазы и транспортируемого вещества. Так, перенос определенного углевода протекает только при участии одной пермеазы. Установлена зависимость поступления в клетки дрожжей сахаров от их циклического строения. Потеря циклического строения у сорбита, маннита и других шестиатомных спиртов приводит к четкому изменению проницаемости. Обнаружена видовая специфичность проникновения сахаров. Клетки Sacch. cerevisiae используют трегалозу, тогда как Sacch. fragilis — нет. Проницаемость клеток в смесях сахаров обусловлена конкуренцией, например, между глюкозой и галактозой, галактозой и мальтозой. Таким образом, физиологическую разнокачественность микроорганизмов определяет не только комплекс ферментов, но и обладание специфической проницаемостью или транспортным механизмом. На процессы переноса растворенных веществ у микроорганизмов влияют факторы окружающей среды Активность пермеаз обычно ингибируется ионами тяжелых металлов, величиной pH, температурой и др. Изменяется проницаемость мембран дрожжей и от условий культивирования. Так, на среде с большим недостатком биотина проницаемость мембран увеличивается [77].
Как и во всех живых организмах, основную часть дрожжевой клетки составляет вода — в пределах 75% от общей массы. Состав сухой массы дрожжей следующий [154], %:
Неорганические вещества 5—10
Белки (N * 6,25) 30—75
Неорганические вещества дрожжевой клетки в основном состоят из фосфорной кислоты (около 50%) и калия (около 25%). Остальные элементы (сера, кальций, железо, хлор, марганец, цинк, молибден, бор и др.) содержатся в ней в незначительных количествах. Углеводы дрожжей составляют полисахариды, гликоген. Содержание свободных аминокислот в дрожжах к концу брожения составляет (в мг/г лиофильно-высушенных дрожжей) [135]: лизин — 7,5; аргинин—1,3; гистидин—11,0; аспарагиновая кислота — 2,9; серии — 2,7; глицин— 1,5; глутаминовая кислота — 3,9; аланин — 8,7; пролин — 2,0; тирозин — 2,8; метионин — 2,9; лейцин (изолейцин) — 5,4; цистеин — следы.
Нуклеиновые кислоты дрожжей — пуриновые и пиримидиновые основания — составляют соответственно 8 и 4% от общего количества азота [154].
Витамины, содержание в 1 г сухих дрожжей, мкг
Инозит 6000— 15000
Пантотеновая кислота 2,0—19,0
Химический состав дрожжей может изменяться в зависимости от состава питательной среды, возраста культуры и условий культивирования. Отношение дрожжей к веществам, входящим в состав среды, зависит главным образом от ферментов, вырабатываемых данным видом или расой дрожжей.
Среды для культивирования дрожжей должны содержать все необходимые им химические элементы и в достаточно легко усвояемой форме,
Углеродное питание. Источниками углерода для дрожжей могут быть самые разнообразные органические соединения углеводы (сахара и их производные), спирты, органические кислоты, аминокислоты, белки, углеводороды и многие другие. Однако в отношении сахаров существует видовая специфичность. На этом построена диагностика видов дрожжей. Так, при общности химизма углеводного обмена большая часть видов рода Saccharomyces различается между собой прежде всего по отношению к сахарам. Что касается других источников углерода — спиртов и органических кислот,— то отношение к ним одинаково у всех видов данного рода [90].
Однако большинство видов винных дрожжей сбраживают глюкозу, фруктозу, мальтозу, сахарозу и галактозу; рафинозу используют частично, а лактозу, мелибиозу, пентозы, декстрины и крахмал совсем не сбраживают. В виноградном сусле примерно в равных количествах содержатся глюкоза и фруктоза. Фруктоза значительно слаще глюкозы, поэтому для приготовления вин с остаточным сахаром лучше использовать дрожжи, обладающие способностью в первую очередь сбраживать глюкозу.
По интенсивности утилизации глюкозы или фруктозы (к моменту, когда сброжено около 50% фруктозы) дрожжи разделяют на 3 группы [154]:
1) глюкозофильные — сбраживают к этому моменту от 80 до 85% глюкозы (большая часть видов рода Saccharomyces, а также виды родов Saccharomycodes и Brettanomyces);
2) фруктозофильные — в этот период используют только от 5 до 10% глюкозы (Sacch. bailli, Sacch. rouxii, T. stellata);
3) дрожжи, использующие оба сахара почти с одинаковой скоростью: к моменту, когда они утилизируют половину фруктозы, исчезает 40—60% глюкозы (Sacch. rosei, Pichia membra-naefaciens).
Органические кислоты занимают важное место в обмене веществ у дрожжей: они могут стимулировать или ингибировать их рост, служить единственным источником углерода и энергии [26,126,178].
Все промежуточные продукты цикла Кребса (пировиноград-ную, уксусную, янтарную, фумаровую и яблочную кислоты) дрожжи в состоянии использовать в качестве единственного источника углерода. Однако скорость роста на средах с этими кислотами ниже, чем на средах с глюкозой. При выдержке вина под хересной пленкой показано образование щавелевой, гликолевой, фумаровой и глутаровой кислот, которых в исходном вине не было [178]. Специальные опыты подтвердили синтез хересными дрожжами из яблочной кислоты фумаровой, янтарной, гликолевой; из пировиноградной — лимонной, яблочной, молочной, янтарной кислот.
Ненасыщенные жирные кислоты, особенно олеиновая, линоле-вая, линоленовая, пальмитолеиновая, арахидииовая, являются важными ростовыми факторами дрожжей в анаэробных условиях. А. Андреазен и Т. Стийер [232] установили, что винные дрожжи могут свободно размножаться в анаэробных условиях, если в среду для культивирования ввести два вещества: какую-либо ненасыщенную жирную кислоту (олеиновую, линолевую, лино-леновую) и стерины (эргостерин или холестерин). П. Брешо с соавторами [239, 242] изучали дрожжи в процессе приготовления вин углекислотной мацерацией. Ими установлено присутствие стимуляторов роста дрожжей в виноградном сусле (пруин винограда), необходимых для развития в анаэробных условиях.
Использование дрожжами углеводородов как единственного источника углерода широко применяется для получения кормового белка. Парафины могут активно потребляться углеводо-родокисляющими дрожжами рода Candida при технологическом процессе производства дрожжевой массы.
В настоящее время существует подкрепленное экспериментальными данными мнение об участии СОг в обмене веществ дрожжей [35]. А. К. Родопуло [161] была доказана способность дрожжей синтезировать из пировиноградной кислоты и углекислоты ряд органических кислот: лимонную, яблочную, янтарную, молочную и др.
При производстве шампанских вин дрожжи оказываются в среде с повышенным содержанием СОг. Л. В. Дубинчук, Н. Н. Глонина, Е. С. Дрбоглав исследовали фиксацию С1402 дрожжами при шампанизации и установили активное использование С02 для синтеза белковых веществ, кислот [54, 56].
Таким образом, среди всех разнообразных органических источников углерода углекислый газ является биологически активным веществом, связанные формы которого являются необходимым продуктом для дрожжей [50].
Азотное питание. Источники азота, необходимые для синтеза азотсодержащих компонентов клетки (аминокислот, белков, пуриновых и пиримидиновых нуклеотидов и некоторых витаминов), должны содержаться в среде в виде органических или неорганических соединений. Большинство дрожжей не усваивают нитраты. Однако род Hansenula характеризуется способностью использовать их, чем и отличается от рода Pichia. Некоторые виды рода Brettanomyces также усваивают нитраты. В качестве неорганических источников азота дрожжи хорошо используют: сернокислый и фосфорнокислый аммоний, аммиачные соли уксусной, молочной, яблочной и янтарной кислот [76].
В этом случае ближайшим предшественником органического азота является аммиак, который дрожжи рода Saccharomyces усваивают в первую очередь и только потом — органические азотистые вещества — аминокислоты. Дрожжи могут использовать в качестве источника азота мочевину и пептон. Для получения большой биомассы Sacch. cerevisiae в аэробных условиях в среде должен содержаться азот как в органической, так и неорганической форме. Один миллиард клеток усваивает около
Аммиачный азот, содержащийся в виноградном сусле (от 25 до 100 мг/л), быстро усваивается дрожжами в первые часы (сутки) размножения клеток. Иногда, при недостатке аммиачного азота в сусле (в годы сильного вызревания винограда или поражения ягод грибом Botrytis cinerea), для усиления роста культуры дрожжей вводят соли аммония. Но делать это можно только перед забраживанием, так как в процессе брожения дрожжи усваивают аммонийные соли не полностью [154]. Следует также обратить внимание на то, что добавки солей аммония повышают в вине титруемую кислотность и снижают величину pH.
Н. Ф. Саенко и другие исследователи установили, что хересные дрожжи лучше развиваются при внесении дополнительных источников азотистого питания (0,5% дрожжевого автолизата или лебедевского мацерационного сока или водного раствора аммиака в количестве 80—120 мг/л). Наиболее быстрому развитию хересной пленки способствует одновременное внесение 0,5% автолизата и 80 мг/л аммиачного азота [178].
Хорошо усваиваются дрожжами аминокислоты, хуже — пептиды и совсем не усваивается нативный белок. Однако при наличии усвояемого азота в среде дрожжи способны расщеплять белок, выделяя протеолитические ферменты [229].
По питательной ценности для вийных дрожжей Э. Пейно и С. Лафон-Лафуркад [292] аминокислоты разделили на хорошо усваиваемые — изолейцин, триптофан, аргинин, валин, гистидин, аспарагиновая кислота и плохо усвояемые — треонин, фенилаланин, тирозин, метионин, серин, лизин, глицин, глутаминовая кислота, лейцин. Совершенно не усваивается пролин.
Примечательным фактом является то, что дрожжи используют только природные формы аминокислот (L-формы).
Дрожжи в процессе брожения виноградного сусла, с одной стороны, потребляют азотистые вещества, с другой — выделяют их в среду. При этом поступление аминокислот и других азотистых веществ значительно увеличивается к концу брожения, когда возрастает число мертвых клеток дрожжей и соответственно — автолитические процессы усиливаются. Однако надо учесть и то, что аминокислоты могут выделяться в среду и живыми клетками.
Н. М. Сисакян и Э. Н. Безингер [188] исследовали изменение состава аминокислот в процессе брожения виноградного сусла и формирования вина и показали, что в процессе сбраживания сусла дрожжи интенсивно ассимилируют большинство аминокислот, не затрагивая пролин, и по окончании брожения отдают в вино аспарагиновую, глутаминовую и у-аминомасляную кислоты, аланин, валин, гликокол, серин, треонин. Таким образом, при культивировании дрожжевых клеток на среде, содержащей полную смесь аминокислот, возможна прямая их ассимиляция в таком количестве, которое соответствует их содержанию в белке дрожжей [76]. Это свидетельствует о том, что для построения белков при размножении и росте клеток необходим не только азот, но и углеродистый остаток аминокислоты. Установлено, что дезаминированный остаток аминокислоты является фактором, определяющим его питательную ценность, позволяет рассматривать аминокислоты не только как источник азота, но одновременно и как источник углерода. Этим можно объяснить принципиальную разницу в ценности различных аминокислот.
Основой метаболизма аминокислот являются реакции трех главных типов: дезаминирование, переаминирование, декарбок-силирование.
В процессе распада и образования «вторичных» аминокислот исключительная роль принадлежит реакциям переаминирования при участии аминотрансфераз. Наличие систем переаминирования у винных дрожжей Sacch. vini и Sacch. oviformis показано В. К. Липатовой [41]. Дрожжи, выращенные в аэробных условиях (виноградное сусло-агар), имели незначительную активность L-аспартат- и L-аланин-аминотрансфераз. У них отсутствовала L-тирозин-аминотрансфераза по сравнению с этими же дрожжами, выращенными в анаэробных условиях (виноградное сусло), хотя выход клеточного экстракта из 1 г сухих дрожжей оставался в тех же пределах. Активность аминотрансфераз дрожжей, выращенных на виноградном сусле, характеризуется данными, приведенными в табл. 9. Несмотря на то что содержание белка в дрожжах рас Киевская и Херес 20 С/96 в аэробных условиях было выше, чем в анаэробных, активность аминотрансфераз была незначительной. Это указывает на связь реакций переаминирования с функциональной деятельностью дрожжей — брожением. Наряду с влиянием условий культивирования на активность аминотрансферазных систем дрожжей оказывал вли-