
Эмбриональное развитие
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
Дробление зиготы
После того, как произошло оплодотворение — слияние сперматозоида и яйцеклетки, образовавшаяся зигота начинает интенсивно делиться. Ее множественные митотические деления называют дроблением.
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
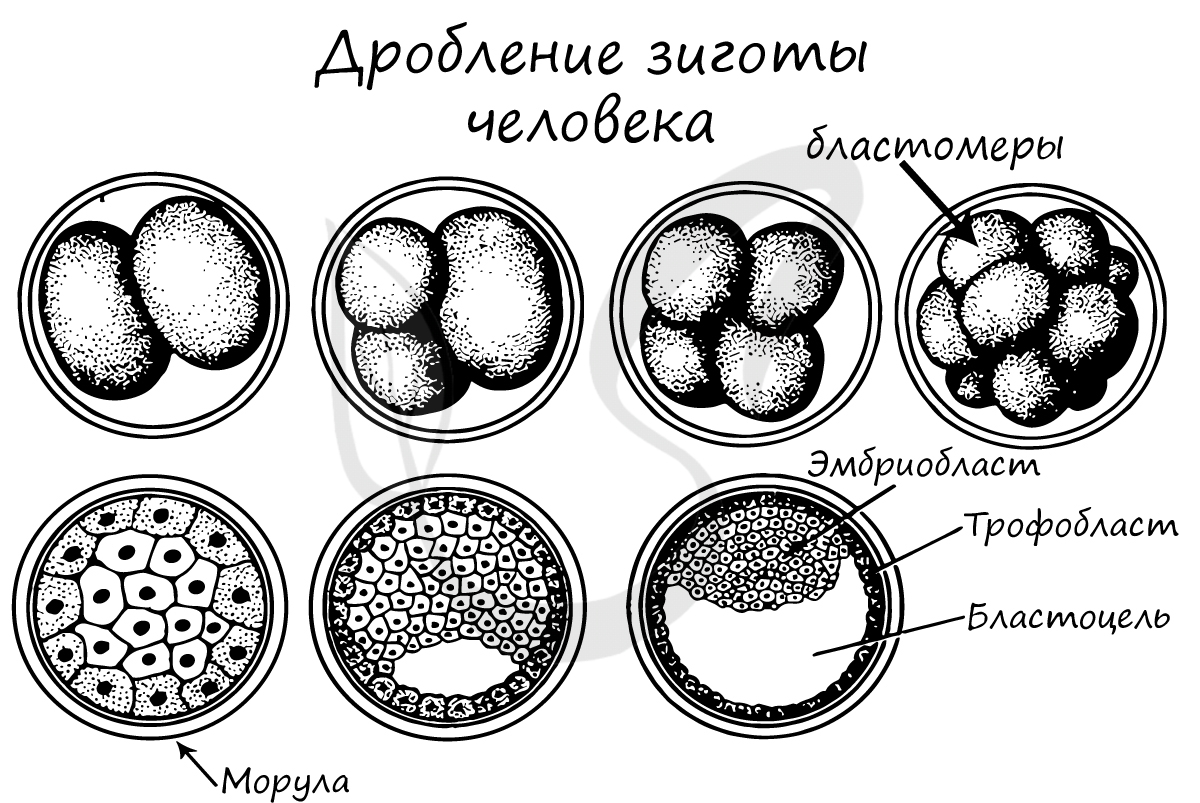
В результате дробления образуется морула. Морула (лат. morum — ягода тутового дерева) — клетка на стадии этапа дробления, когда зародыш представляет собой компактную совокупность клеток (без полости внутри).
Бластуляция
Бластуляция — заключительный период дробления, в который зародыш называется бластулой.
После очередных этапов многократного деления образуется однослойный зародыш с полостью внутри — бластула (греч. blastos — зачаток).
Стенки бластулы состоят из бластомеров, которые окружают центральную полость — бластоцель (греч. koilos — полый). Соединяясь друг с другом, бластомеры образуют бластодерму из одного слоя клеток.

Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.
Стенка бластулы начинается впячиваться внутрь — происходит инвагинация стенки. По итогу такого впячивания зародыш становится двухслойным. Двухслойный зародыш называется — гаструла. Полость гаструлы называется гастроцель (полость первичной кишки), а отверстие, соединяющее гастроцель и внешнюю среду — первичный рот (бластопор).

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).

При впячивании части бластулы (инвагинации) клетки бластодермы мигрируют внутрь и становятся энтодермой (греч. entós — внутренний). Оставшаяся часть бластодермы снаружи называется эктодермой (греч. ἔκτος — наружный).
Между энто- и эктодермой из группы клеток формируется третий зародышевый листок — мезодерма (греч. μέσος — средний).

Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.
Важно отметить, что на стадии нейрулы происходит процесс нейруляции — закладывание нервной трубки. Нервная пластинка, образовавшаяся на ранних этапах, прогибается внутрь, при этом ее края сближаются и, замыкаясь, формируют нервную трубку.

Итак, как уже было сказано, на стадии нейрулы закладываются отдельные органы. Эктодерма образует покровный эпителий и нервную пластинку, мезодерма (из которой в дальнейшем появятся все соединительные ткани), энтодерма — окружает полость первичной кишки (гастроцель), образуя кишечник. От энтодермы отшнуровывается хорда.
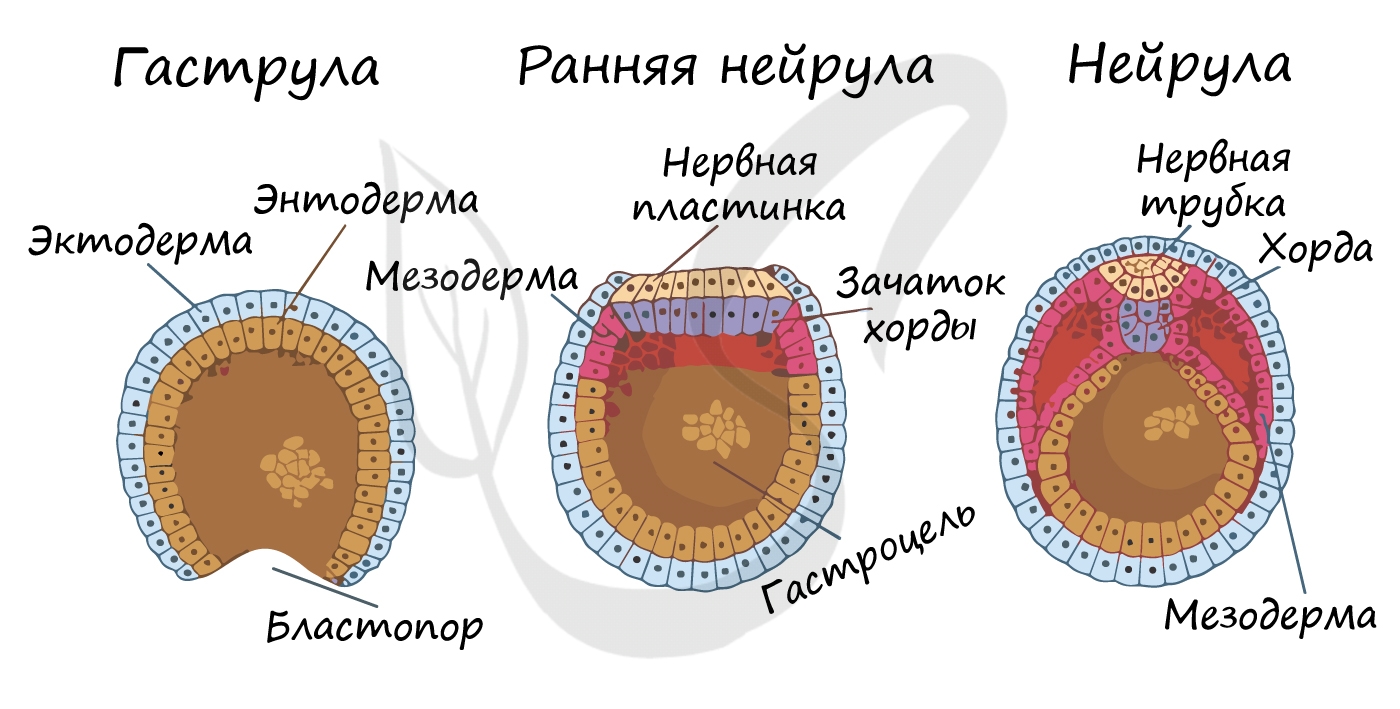
Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.
Эктодерма (греч. ἔκτος — наружный) — наружный зародышевый листок, образует головной и спинной мозг, органы чувств, периферические нервы, эпителий кожи, эмаль зубов, эпителий ротовой полости, эпителий промежуточного и анального отделов прямой кишки, гипофиз, гипоталамус.
Мезодерма (греч. μέσος — средний) — средний зародышевый листок, образует соединительные ткани: кровеносную и лимфатическую системы, костную и хрящевую ткань, мышечные ткани, дентин и цемент зубов, а также выделительную (почки) и половую системы (семенники, яичники).
Энтодерма (греч. entós — «внутренний») — внутренний зародышевый листок, образует эпителий пищевода, желудка, кишечника, трахеи, бронхов, легких, желчного пузыря, мочевого пузыря и мочеиспускательного канала, печень и поджелудочную железу, щитовидную и паращитовидную железы.

Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
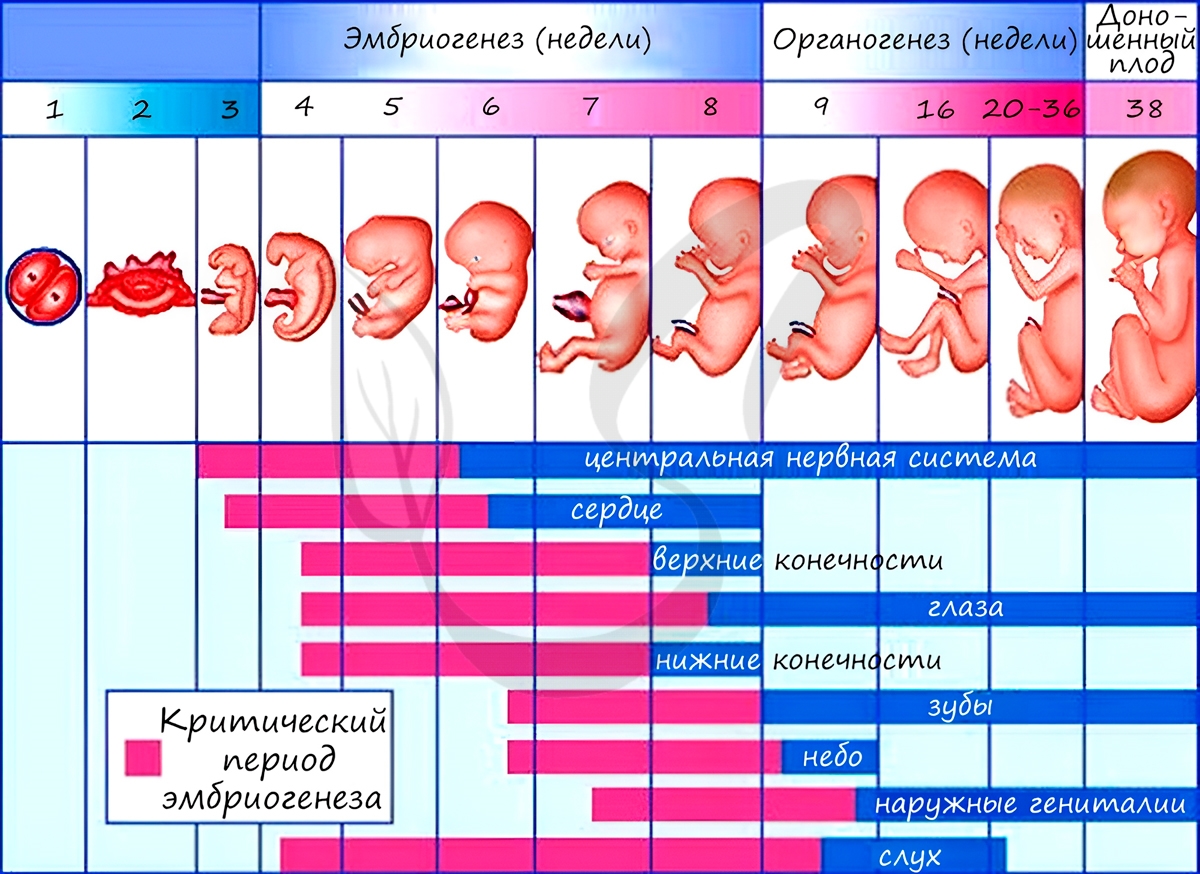
Анамнии и амниоты
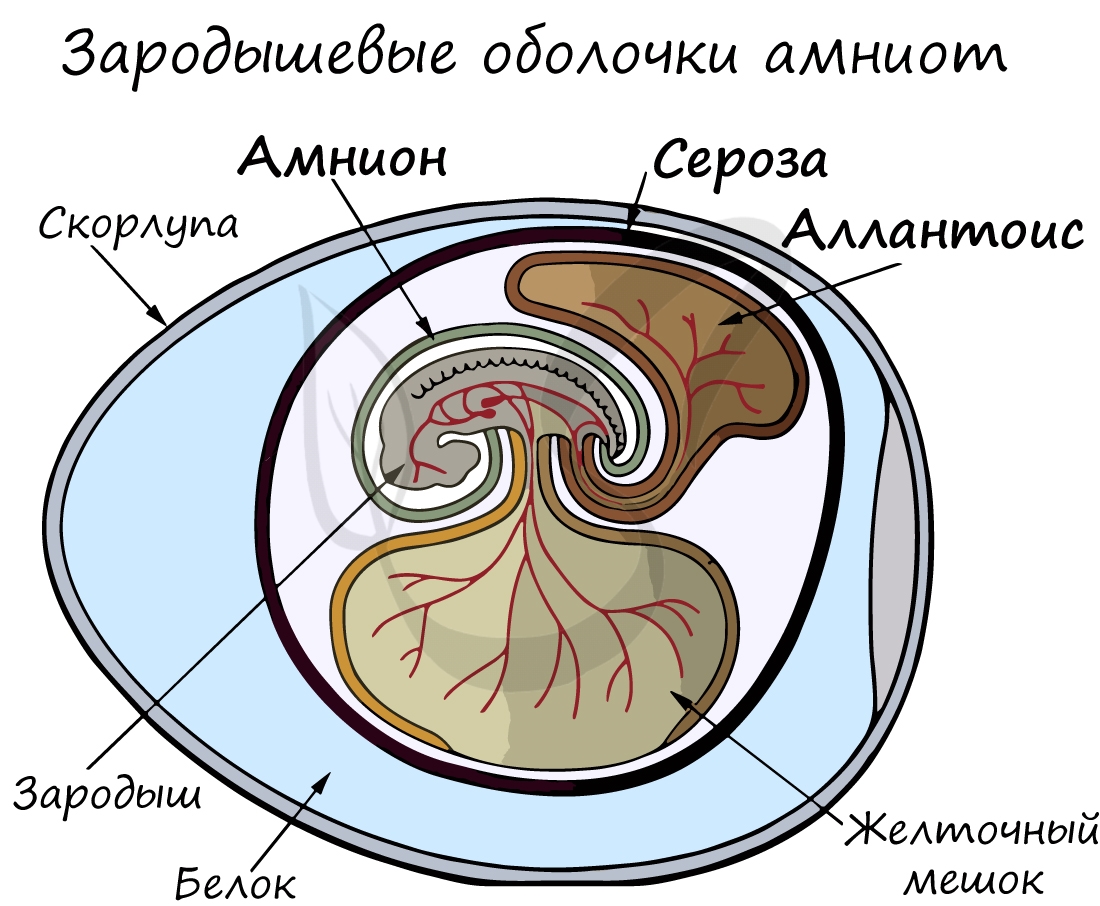
Анамнии, или низшие позвоночные — группа животных, не имеющая зародышевых оболочек (зародышевого органа — аллантоиса и амниона). Анамнии проводят большую часть жизни в воде, без которой невозможно их размножение.
К анамниям относятся рыбы, земноводные.

Амниоты — группа высших позвоночных, характеризующаяся наличием зародышевых оболочек. К амниотам относятся пресмыкающиеся, птицы и млекопитающие.
Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они «обрели независимость» от него.
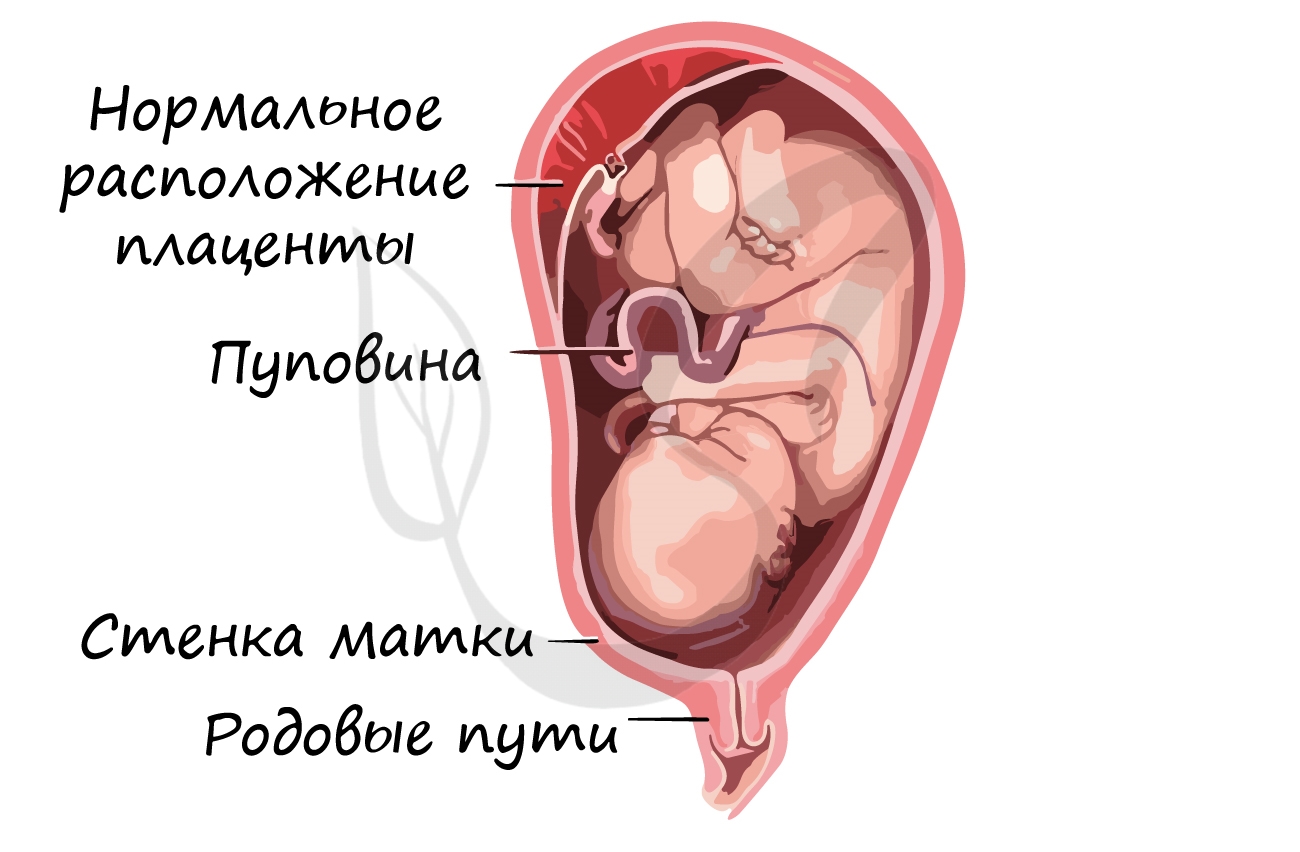
Развитие плода происходит в мышечном органе — матке, которая, сокращаясь во время родов, стимулирует изгнание плода через родовые пути. Питание осуществляется через плаценту — «детское место» — орган, который с одной стороны омывается кровью матери, а с другой — кровью плода. Через плаценту происходит транспорт питательных веществ и газообмен.
Соединяет плаценту и плод особый орган — пуповина, внутри которой проходят артерии, вены.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Источник
Классификация дробление и яиц.
Яйца классифицируют следующим образом:
1. По количеству желтка — полицитальные, мезоцитальные, олигоцитальные и алецитальные (т.е. многожелтковые, среднежелтковые, маложелтковые и безжелтковые).
2. По распределению желтка по объему яйца:
— телолецитальные — количество желтка возрастает от аномального полюса к вегетативному;
— гомолецитальные (изолецитальные) — желточные гранулы равномерно распределены в толще яйца;
— центролецитальные — свободная от желточных гранул цитоплазма расположена сразу под оболочкой яйца, вокруг ядра, занимающего центральное положение, и в виде тонких тяжей, соединяющих названные области. Промежуточное пространство заполнено желтком. (Кноррэ А. Г)
Классификация типов дробления зиготы
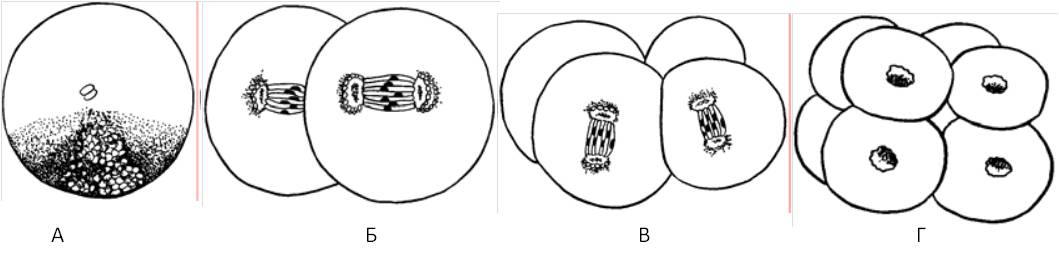
Голобластический тип — полное разделение яйца и бластомеров бороздами дробления (а-, олиго-, мезолецитальные, изо-, телолецитальные).

Голобластическое дробление иглокожих (по Гилберту, 1993):
А, Б, В – последовательные стадии процесса
2. Меробластический тип — частичное разделение яйца. Борозды дробления проникают в глубь яйца, но не разделяют его полностью. Желток остается неразделенным:
— поверхностное дробление (полилецитальные, центролецитальные яйцеклетки) — разделение поверхностного слоя цитоплазмы с одиночными (предварительно многократно поделившимися) ядрами посредством перегородок, направленных нормально к поверхности яйца. Центральная часть яйца остается не разделившейся:
— дискоидальное дробление (полилецитальные, телолецитальные яйцеклетки) — борозды формируются вслед за делениями ядер, но не разделяют все яйцо, а только один из его полюсов.

Дискоидальное дробление куриного яйца (по Гилберту, 1993):
Представлен вид со стороны анимального полюса. Борозды дробления не распространяются на желток, а возникающая бластодерма состоит из одного слоя клеток
3. По признаку объемов, образующихся в результате дробления:
— равномерное — объемы бластомеров одинаковы:
— неравномерное — объемы бластомеров неодинаковы.
4. По признаку продолжительности карио- и цитотомии в разных бластомерах дробящегося яйца:
— синхронное — дробление начинается и завершается во всех бластомерах одновременно;
— асинхронное — начало и время деления в разных бластомерах неодинаково.
5. По признаку взаимного расположения бластомеров в дробящемся яйце:
— радиальное дробление — взаимное расположение бластомеров таково, что исходная полярная ось яйца служит осью радиальной симметрии дробящегося зародыша;
— спиральное дробление — прогрессивное нарушение симметрии дробящегося яйца в результате спирального смещения завершающих деление бластомеров относительно друг друга;
— билатеральное дробление — бластомеры расположены так, что через зародыш можно провести только одну плоскость симметрии;
— анархическое дробление — отсутствие закономерности в расположении бластомеров.

Анархическое дробление (по Токину, 1987)
Дробление завершается образованием бластулы — многоклеточной структуры с более или менее выраженной полостью внутри. Последняя называется полостью дробления, или бластоцелем. (Карлсон Б. М)
Дробление у ланцетника.
Малое количество желтка объясняет простоту дробления и гаструляции. Дробление полное, почти равномерное, радиального типа, в результате образуется целобластула.
Дробление яйца ланцетника (по Алмазову, Сутулову, 1978):
А – зигота; Б, В, Г – образование бластомеров
(показано расположение веретена деления)
Анимальный полюс приблизительно соответствует будущему переднему концу тела личинки. Оплодотворенная яйцеклетка (зигота) целиком дробится на бластомеры в правильной геометрической прогрессии. Бластомеры почти одинаковой величины, анимальные лишь несколько мельче вегетативных. Первая борозда дробления – меридинальная, проходит через анимальный и вегетативный полюс. Она разделяет шаровидное яйцо на две совершенно симметричные половины, но бластомеры округляются. Они шаровидные, имеют малую площадь соприкосновения. Вторая борозда дробления также меридиональная, пер-
пендикулярна первой, а третья – широтная.
По мере увеличения количества бластомеров они все больше расходятся от центра зародыша, образуя посередине большую полость. В конце концов зародыш принимает форму типичной целобластулы – пузырька со стенкой, образованной одним слоем клеток – бластодермой и с по-
лостью, заполненной жидкостью – бластоцелем.
Клетки бластулы, вначале округлые и потому не плотно сомкнутые, затем приобретают форму призм и плотно смыкаются. Поэтому позднюю бластулу, в противоположность ранней, называют эпителиальной. Стадия поздней бластулы завершает период дробления. К концу этого периода размеры клеток достигают минимума, а общая масса зародыша не увеличивается по сравнению с массой оплодотворенной яйцеклетки.
Дробление у амфибий.
Дробление у амфибий голобластическое (полное), неравномерное и асинхронное. Первая борозда дробления меридиональная, проходит отанимального полюса к вегетативному. Поскольку у амфибий вегетативный полюс перегружен желтком, то чем ближе к нему, тем скорость врезания борозды становится меньшей. Она еще не успевает дойти до вегетативного полюса, как начинает врезаться 2-я борозда.
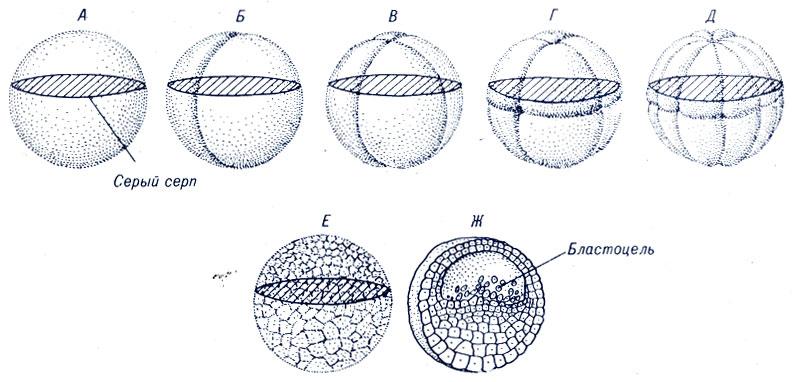
Рис. 3. Дробление (А – Е) зиготы и поперечный разрез бластулы (Ж) лягушки
(по Гилберту, 1993)
Вторая борозда тоже меридиональная, проходит перпендикулярно первой. После прохождения первой борозды образуется два, после второй – четыре бластомера. На стадии 4-х бластомеров два анимальных бластомера получают по ½ материала серого серпа, а в 2-х вегетативных бластомерах материала серого серпа нет.
Третья борозда дробления широтная, проходит ближе к анимальному полюсу. В результате ее прохождения образуются 8 бластомеров: 4 анимальных и 4 вегетативных, существенно различающихся своими размерами. Далее следуют две меридиональные борозды. Из-за разницы в размерах бластомеров и количества желтка вначале делятся 4 анимальных (кратковременная стадия 12 бластомеров), а затем 4 вегетативных бластомера (стадия 16 бластомеров).
Следующие две одновременно возникающие борозды — широтные. Более быстрое прохождение верхней широтной борозды обусловливает 6 кратковременную стадию 24 бластомеров. Завершение прохождения нижней широтной борозды приводит к образованию 32 бластомеров. После стадии 64 бластомеров несмотря на то, что последовательность борозд сохраняется, геометрическая последовательность ряда (2 — 4 — 8 — 12 — 16 — 24 — 32 — 64) нарушается. Параллельно с прохождением меридиональных и широтных борозд дробления образуется и тангенциальная борозда, в результате чего зародыш приобретает многослойность.
Зародыш амфибий, содержащий от 16 до 64 клеток, обычно называют морулой из-за отдаленного внешнего сходства с тутовой ягодой (лат. morum). На 128-клеточной стадии появляется хорошо различимый бластоцель и принято считать, что в это время зародыш достигает стадии бластулы, хотя образование бластоцеля прослеживается с самого первого деления дробления.( Голиченков В.А.)
Дробление у рыб
Яйца полилецитальные, 
полиспермия. Полагают, что
ядра спермиев, не слившихся с
женским пронуклеусом, долго
сохраняются в желточном слое
яйца, граничащем с бластодермой,
и даже участвуют в переработке
желтка. Дробление дискоидальное.
Пятое деление дробления
отделяет ряд бластомеров от
желтка. Краевые бластомеры
и бластомеры, лежащие в основании бластодермы, сохраняют связь с желтком. Шесть делений дробления проходят более или менее синхронно, после чего синхронность нарушается. В итоге в основании бластодермы формируется перибласт или слой мероцитов. В результате дробления образуется дискобластула, снаружи ограниченная слоем плотно соединенных покровных клеток. Со временем щелевидная полость бластулы увеличивается, причем в большей степени у одного из краев бластодиска. Полагают, что различия в размере бластоцеля (или, иными словами, плотность упаковки клеток в различных частях бластодермы) определяет оси и билатеральную симметрию будущего зародыша — область расширенного бластоцеля соответствует каудальному концу зародыша.
Дробление у пресмыкающихся
Дробление яйцеклетки у пресмыкающихся неполное, дискоидальное. Так как оплодотворение яйцеклетки происходит в верхней трети яйцевода, то дробление начинается во время движения яйцеклетки по яйцеводу и при откладывании яйца зародыш находится либо на стадии дискобластулы, либо ранней гаструлы. У ряда пресмыкающихся, в связи с длительным нахождением яйца в яйцеводе происходит формирование нового организма, который рождается, или выходит из яйца сразу же после его откладывания (живородящая ящерица, гадюки).
Дискоидальное дробление приводит к возникновению дискобластулы. Крыша бластулы состоит из мелких клеток бластодермы, совокупность которых называется бластодиском, а дно бластулы образовано нераздробленной массой желтка. Часть желтка под зародышевым диском резорбируется и образуется подзародышевая полость в виде щели между внешним и внутренним слоем бластодиска.
Дробление у птиц
Дробление и формирование бластулы у птиц проходят при движении яйца вниз по яйцеводу, т.е. одновременно с окружением его третичными оболочками. Период дробления у куриного зародыша составляет 22 часа.
Дробление зиготы у птиц неполное, дискоидального типа. Дроблению подвергается только зародышевый диск, расположенный на анимальном полюсе, имеющий ничтожные поверхность и объем по сравнению с недробящейся массой желтка. Первые три борозды дробления радиальные, соответстветствующие меридиональным бороздам ланцетника и амфибий, затем возникают широтные и тангенциальные борозды (рис. 2).

Рис. 2. Дискоидальное дробление у курицы (по Дондуа, 2005):
А – 2 бластомера; Б – 4 бластомера; В – 8 бластомеров; Г – 16 бластомеров
При дискоидальном дроблении на краю бластодиска и под ним все время остаются неполностью отделенные от желтка клетки (перибласт), от которых обособлены центральные бластомеры. Дробление у птиц резко неравномерное и неправильное. Начиная с прохождения третьей и четвертой борозд образуются клетки разной величины, без какой-либо правильности и постоянства в их расположении.
К моменту откладки яйцо проходит около 14 делений зиготы, в результате бластодерма только что отложенного яйца насчитывает около 60 тыс. клеток. В итоге образуется многослойный бластодиск, состоящий из клеток неправильной формы, вплотную прилежащих друг к другу и налегающих на нераздробившийся желток (рис. 3).

Рис. 3. Формирование зародышевого диска курицы (по Дондуа, 2005):
А – поздняя стадия дробления; Б – перед началом гаструляции:
1 – бластомеры; 2 – эпибласт; 3 – подзародышевая полость; 4 – желток
Под влиянием клеток бластодиска и мероцитов часть желтка под зародышевым диском разжижается, образуется небольшое пространство,
заполненное жидкостью – подзародышевая полость. На этой стадии четко различимы две области бластодиска: светлое поле в центре (area pellucida) и темное по периферии (area opaca). (Голиченков, В. А. Практикум по)
Появление светлого поля связано с тем, что в центре бластодиска
зародышем раньше всего используется желток, вследствие чего образуется подзародышевая щель и вся эта область выглядит прозрачной. Клетки периферической зоны по краю бластодиска налегают на желток, поэтому этот участок зародыша выглядит темным.( Голиченков, В. А)
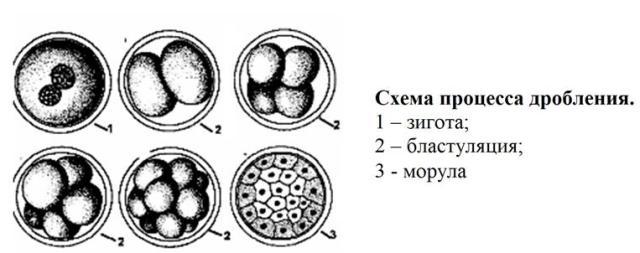
Дробление умлекопитающих. Вскоре после образования зиготы начинается ряд митотических делений, строго контролируемых геномом и называемых дроблением; этот процесс начинается спустя несколько часов после оплодотворения, еще в яйцеводах. Каждое деление продолжается от 12 до 24 часов. Зигота делится на клетки-бластомеры в определенной последовательности. Первое деление происходит в плоскости, проходящей через оба полюса яйца, таким образом, шаровидная зигота делится на два полушаровидных бластомера. Возникающая борозда деления располагается не случайным образом, а в зависимости от места проникновения спермия и последующего перераспределения цитоплазмы. Вторая борозда дробления проходит перпендикулярно предыдущей. Получаются четыре бластомера, группирующихся по принципу радиальной симметрии, затем они снова делятся пополам с образованием восьми бластомеров. Продолжающиеся деления бластомеров характеризуются чередованием плоскостей деления – один из двух бластомеров делится в экваториальной плоскости, другой – в меридиональной. Такой тип деления, типичный для млекопитающих, называется чередующимся. Причем бластомеры делятся не одновременно, и в результате этого не происходит четкого нарастания числа клеток от 2 к 4, а далее к 8. В определенные моменты времени зародыши содержат нечетное число бластомеров. Каждая из клеток, образовавшихся при дроблении, меньше материнской примерно в 2 раза. В период между делениями дробления не происходит интерфазного роста клеток, и поэтому общая масса всех клеток остается примерно равной массе яйцеклетки.  Рис. 1. Ранние стадии эмбрионального развития у млекопитающих:
Рис. 1. Ранние стадии эмбрионального развития у млекопитающих:
I – начало дробления; А – оплодотворения; В, С, D – бластомеры; II – образование морулы — постепенное обрастание темных клеток светлыми; III – образование бластоцисты: А – начало образования полости; В – обособление зародышевого узелка от трофобласта; C – превращение зародышевого узелка в зародышевый щиток (дискобластула); 1 – трофобласт; 2 – эмбриобласт. В результате делений формируется многоклеточный зародыш, напоминающий по внешнему виду ягоду малины и называемый морулой. На стадии 8-клеточной морулы, что характерно только для млекопитающих, происходит значительное сближение 6ластомеров. При этом между клетками возникают плотные контакты, позволяющие некрупным молекулам и ионам переходить из клетки в клетку. Это явление носит название компактизации и способствует дальнейшей дифференцировке зародыша и отделению трофобласта, берущего на себя питательную функцию, и внутренней клеточной массы, дающей начало собственно эмбриону. В процессе компактизации отдельные части плазматической мембраны клеток зародыша начинают двигаться в разных, строго генетически детерминированных направлениях. В этих процессах участвуют белки, входящие в состав клеточных мембран, и с возникновением микроворсинок, прикрепляющих бластомеры друг к другу, изменяется их цитоскелет. На стадии 32-клеточной морулы зародыш собаки попадает в матку. Это происходит на 7-8-й день, что значительно позже, чем у других млекопитающих. За это время прекращается развитие и происходит гибель зародышей с нарушениями деления, вызванными как наследственными, так и средовыми факторами. Таким образом, наследственные и средовые факторы являются действующими факторами естественного отбора. Бластомеры, получающиеся в процессе деления, располагаются по периферии, и постепенно в середине плотной клеточной массы морулы формируется полость (бластощель) и отделяется собственно зародышевое скопление клеток – эмбриобласт, иначе называемый зародышевым узлом, и окружающий его слой питающих клеток – трофобласт. Из эмбриобласта позднее формируется тело зародыша. Трофобласт служит питающим листком для раннего зародыша. Эта стадия развития зародыша называется бластоцистой. Бластоциста на этом этапе состоит из наружного слоя клеток трофобласта и внутренней клеточной массы, шаровидного скопления клеток, прикрепленного изнутри к одному из полюсов трофобласта и представляющего собой материал для построения зародыша. Будущее расположение клетки в зародыше или трофобласте определяется в процессе компактизации, когда клетки оказываются или на поверхности, или внутри зародыша.
Источник



