
Методы дифференциального окрашивания хромосом




![]()
![]()
В начале 70-х гг. были разработаны методы дифференциальной окраски хромосом, которые позволяли однозначно идентифицировать каждую хромосому.
Методы были основаны на способности некоторых красителей специфически связываться с конкретными участками хромосом в зависимости от их структурно-функциональной организации. Предложенные методы выявляли линейную неоднородность (сегменты) хромосом.
На практике наибольшее применение получили методы дифференциальной окраски красителем Гимза (G-окраска) и флюоресцирующим красителем акрихином или акрихинипритом (Q-окраска) (рис. 6.5, 6.6).
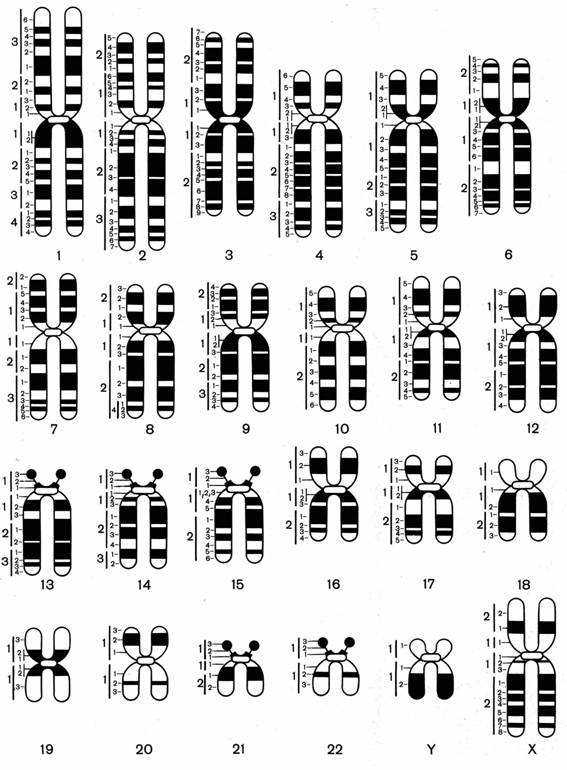
Рис. 6.5. Дифференциальная окраска хромосом
На рис. 5 представлены хромосомы человека при G-окраске. Хорошо видно, что каждая хромосома человека имеет только ей свойственную последовательность разношироких полос. Это позволяет точно идентифицировать любую из хромосом и обнаруживать относительно крупные изменения в их структуре.
При анализе метафазных хромосом средней конденсации можно четко различить около 350-400 относительно крупных сегментов на гаплоидный набор.
На стадиях, предшествующих метафазе, хромосомы менее спирализованы и поэтому имеют большую поперечную подразделенность.
Были разработаны методы анализа хромосом на делящихся клетках в стадии прометафазы. Использование этого методического подхода позволило получить хромосомы с разной степенью сегментации — от 800 до 2500 сегментов на гаплоидный набор.
На рис. 6 представлены дифференциально окрашенные X и Y-хромосомы с различными уровнями спирализации. Использованный подход дает возможность точно устанавливать точки разрывов в перестроенных хромосомах, даже если в перестройку вовлечены небольшие участки хромосом.
Поперечная исчерченность, обнаруживаемая различными методами, в принципе выявляет одни и те же сегменты хромосомы и является результатом неравномерной конденсации хроматина по всей ее длине. В зависимости от степени спирализации ДНК в хромосоме выделяют гетерохроматиновые и эухроматиновыерайоны, для которых характерны различные функциональные и генетические свойства.
Гетерохроматиновый район представляет собой участок конденсированного хроматина (высокоспирализованная ДНК), который выявляется при дифференциальном окрашивании в виде темных полос. Присутствие гетерохроматина можно обнаружить и в интерфазном ядре, где он отчетливо выявляется в виде интенсивно окрашенных глыбок хроматина. Считывания генетической информации с данных участков не происходит.
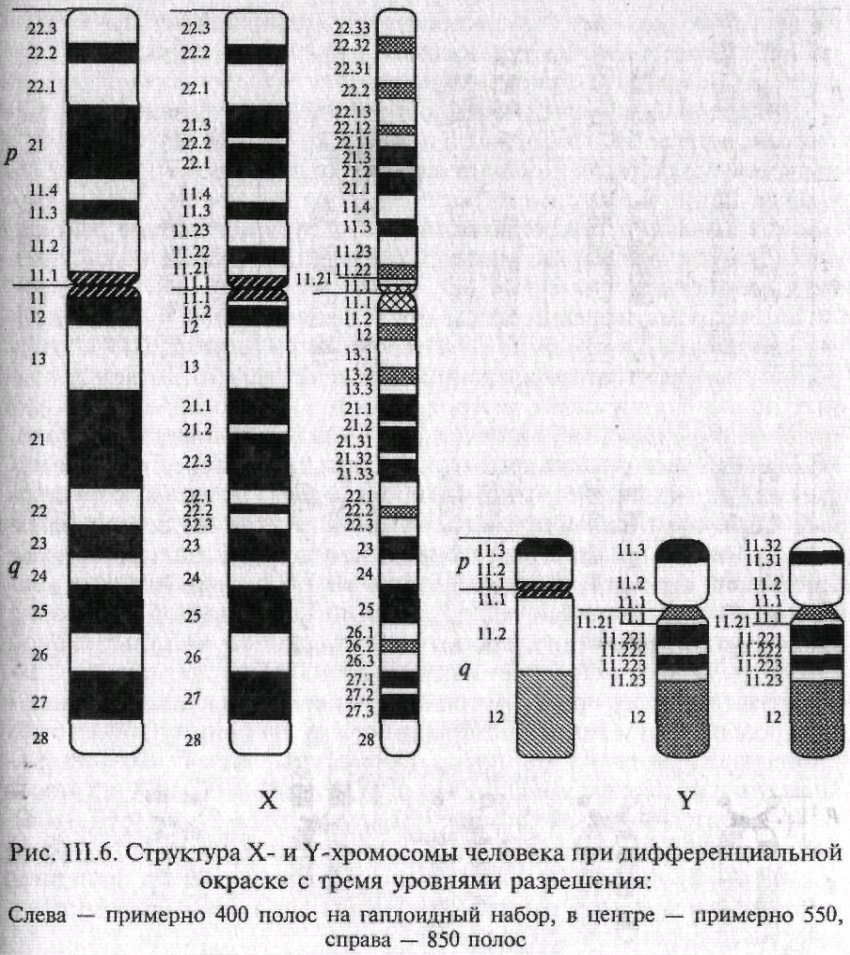
Рис. 6.6. Структура хромосом человека при дифференциальной окраске с тремя уровнями разрешения
Различают структурный и факультативныйгетерохроматин. Структурный гетерохроматин постоянно присутствует в определенных регионах хромосомы. Например, он всегда обнаруживается вокруг центромер всех хромосом. Факультативный гетерохроматин появляется в хромосоме при сверхспирализации эухроматиновых районов. Факультативной гетерохроматизацией может быть охвачена целая хромосома.
Так, в клетках женского организма одна из Х-хромосом полностью инактивирована путем гетерохроматизации уже на ранних этапах эмбрионального развития. Ее можно обнаружить в виде глыбки гетерохроматина на периферии ядра. Такая инактивированная Х-хромосома называется половым хроматином, или тельцем Барра.
Благодаря гетерохроматизации Х-хромосомы в клетках женского организма происходит выравнивание количества генов, функционирующих в мужском и женском организмах, поскольку у мужчин имеется только одна Х-хромосома.
Эухроматиновые регионы хромосом в интерфазном ядре не видны, поскольку представлены хроматином в деконденсированном состоянии. Это указывает на их высокую метаболическую активность. Действительно, эухроматиновые районы содержат уникальные гены, контролирующие синтез различных белков. При дифференциальном окрашивании метафазных хромосом они определяются как светлые полосы.
Изучение химического состава хромосом показало, что они состоят из дезоксирибонуклеиновой кислоты (ДНК), гистонов, негистонных белков и небольшого количества РНК.

В каждой хромосоме содержится только одна молекула ДНК. Однако размеры молекул ДНК хромосом огромны. Они могут достигать сотен микрометров и даже сантиметров. В кариотипе человека самая большая хромосома — 1; длина ее полностью раскрученной ДНК составляет около 7 см, что значительно больше не только размеров ядра клетки, но и самой клетки. Суммарная длина молекул ДНК всех хромосом одной клетки человека составляет более 170 см. Несмотря на свои гигантские размеры, молекулы ДНК функционируют в пределах таких микрообразований, как хромосомы. Поэтому хромосомы ядер клеток должны представлять собой сильно укороченные (конденсированные) структуры ДНК. Это достигается за счет специфической укладки молекул ДНК — многоуровневой спирализации. Основной структурной единицей хромосомы является нуклеосома (рис. 6.7, 6.8).
Каждая нуклеосома содержит по две молекулы четырех различных типов гистонов, объединенных в октамер (восьмигранник), обвитый нитью ДНК. Нуклеосомы и соединяющие их участки ДНК формируют спиральную структуру — хроматиновое волокно. На каждый виток такой спирали приходится 6 нуклеосом. Так формируется структура хромосомы (рис. 6.7, 6.8).
При конденсации происходит уменьшение длины молекулы ДНК в 10 тыс. раз, так что конденсированные хромосомы в среднем имеют длину порядка 200 нм (т. е. 200х10 -9 м). Это обеспечивает возможность точного и быстрого деления генетического материала материнской клетки между дочерними клетками (митоз) и уменьшение числа хромосом вдвое при образовании половых клеток (мейоз).
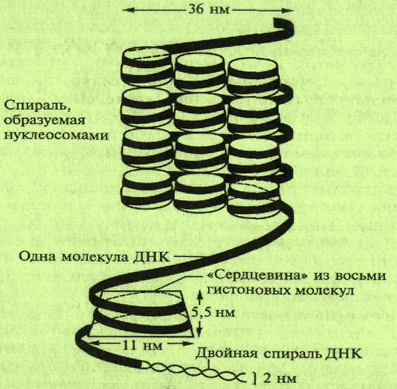
Рис. 6.7. Структура нуклеосомы
Хромосомы выполняют функцию основного генетического аппарата клетки. В них в линейном порядке расположены гены из которых занимает строго определенное место, называемое локусом. Альтернативные формы гена (т.е. различные его состояния), занимающие один и тот же локус, называются аллелями (от греч. allelon — взаимно другой, иной). Любая хромосома содержит только единственный аллель в данном локусе, несмотря на то, что в популяции могут существовать два, три и более аллелей одного гена.
Уровни организации хроматина(рис. 6.8)
• метафазная хромосома – 1400 нм
• участок метафазной хромосомы – 700 нм;
нуклеомеры- субъединицы толстой фибриллы — 300 нм
Источник
Дифференциальная окраска хромосом.
![]()
![]()
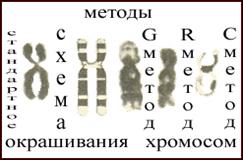
В зависимости от целей цитогенетического исследования используются различные методы окрашивания хромосом (см. рис. 9). Наиболее распространенными из них являются рутинная или обычная окраска и ряд методов дифференциального окрашивания хромосом: Q-, G, С-, R- и NOR- или Ag-окраска.В свою очередь, методы
дифференциального окрашивания делятся на две группы:
1) приводящие к образованию сегментов вдоль длины всех хромосом (например Q-, G- или R-сегменты);
2) приводящие к окрашиванию специфических хромосомных структур, в результате чего выявляется ограниченное число сегментов (С-, Т- или NOR-сегменты). Рис.9.Методы окрашивания хромосом (8)
 Рутинная окраскахромосом достигается путем простого окрашивания полученных хромосомных препаратов красителем Романовского-Гимза (азурэозином) или 2% ацетоорсеином, или 2% ацетокармином, без какой-либо их предварительной обработки. Такая окраска приводит к равномерному и интенсивному прокрашиванию хромосом по длине, что не позволяет идентифицировать разные морфологически сходные хромосомы набора. Исторически рутинная окраска была самой первой используемой в цитогенетике человека окраской, активно применяющейся в течение 1959-1970 гг., до внедрения методов дифференциального окрашивания хромосом. В настоящее время она практически не применяется для диагностики конституциональных хромосомных нарушений, однако находит применение при анализе хромосомных аберраций в тестировании факторов среды на мутагенную активность. Такую окраску можно применять для выявления численных аномалий хромосом.
Рутинная окраскахромосом достигается путем простого окрашивания полученных хромосомных препаратов красителем Романовского-Гимза (азурэозином) или 2% ацетоорсеином, или 2% ацетокармином, без какой-либо их предварительной обработки. Такая окраска приводит к равномерному и интенсивному прокрашиванию хромосом по длине, что не позволяет идентифицировать разные морфологически сходные хромосомы набора. Исторически рутинная окраска была самой первой используемой в цитогенетике человека окраской, активно применяющейся в течение 1959-1970 гг., до внедрения методов дифференциального окрашивания хромосом. В настоящее время она практически не применяется для диагностики конституциональных хромосомных нарушений, однако находит применение при анализе хромосомных аберраций в тестировании факторов среды на мутагенную активность. Такую окраску можно применять для выявления численных аномалий хромосом.
Q-окраска(от англ. Quinacrine — акрихин) выявляется на хромосомах в виде чередования ярко- и темно-флюоресцирующих полос с помощью флуоресцентной микроскопии хромосомных препаратов, окрашенных такими флюорохромами (флуоресцентными красителями) как производные акридина — акрихин дигидрохлорид (атебрин) или акрихин-иприт. Эти красители обладают способностью присоединяться к ДНК путем интеркаляции или с помощью внешних ионных сил. Q-окраска имеет свою кодировку (QFQ)по международной цитогенетической номенклатуре. Заслуга применения производных акрихина для получения Q-сегментов на хромосомах человека принадлежит шведскому цитогенетику Касперссону (1970 г.). В популяциях человека существует межиндивидуальная вариабельность отдельных участков хромосом, выявляемых с помощью Q-окраски — Q-полиморфизм хромосом. Он выражается в особенно ярко светящихся сегментах, локализованных в центромерных участках (сеn) хромосом 3 и 4, а также в коротких плечах (p11) и спутниках (р13) всех акроцентрических хромосом человека 13-15, 21 и 22. Кроме того, у лиц мужского пола ярко флюоресцирующейся областью является и дистальная часть длинного плеча хромосомы Y — сегмент q12. Этот участок Y-хромосомы обладает выраженным межиндивидуальным полиморфизмом и четко наследуется по мужской линии. Все перечисленные ярко флюоресцирующие участки хромосом являются областями локализации некодирующих повторяющихся гетерохроматиновых последовательностей ДНК, вариабельность которых не приводит к каким-либо фенотипическим изменениям. Указанные выше ярко флюоресцирующие Q-полиморфные хромосомные сегменты являются удобными цитогенетическими маркерами и могут быть использованы как для характеристики популяций, индивидов и клеточных линий, так и для решения некоторых судебно-медицинских проблем. Метод позволяет даже в интерфазном ядре идентифицировать У-хромосому человека по яркой флуоресценции.
G-окраска(от англ. Giemsa — Гимза) выявляется благодаря предварительной обработке хромосомных препаратов слабым раствором протеолитического фермента трипсина и последующей окраске красителем Гимза. При этом наблюдается полосатая исчерченность хромосом, где темные полосы в некоторой степени соответствуют гетерохроматиновым районам, а светлые -эухроматиновым. G-окраска имеет свою кодировку (GTG) по международной цитогенетической номенклатуре. Оптимальные условия окраски находят в каждой лаборатории эмпирическим путем. Методика G-окраски хромосом человека была впервые предложена английской исследовательницей М. Сибрайт в 1972 году и практически в неизменном виде используется до настоящего времени. По числу, величине и расположению выявляющихся сегментов рисунок G-окраски аналогичен рисунку при Q-окраске, где темно окрашенные G-сегменты соответствуют флюоресцирующим Q-сегментам. Различия состоят в том, что: а) несветящиеся гетерохроматиновые центромерные сегменты в хромосомах 1 и 16 хорошо прокрашиваются красителем Гимза; б) ярко флюоресцирующие при Q-окраске сегменты 3, 4, 13-15, 21, 22 и Y-хромосом не выделяются особой интенсивностью при G-окраске. На G-окрашенных метафазных хромосомах выделяется около 320 сегментов на гаплоидный геном. Преимущество метода в том, что он не требует использования флуоресцентного микроскопа и окрашенные препараты можно длительно хранить.
R-окраска(от англ. Reverse — обратная) отличается противоположностью рисунка G-окраске. Темноокрашенными здесь являются эухроматиновые участки хромосом, а светлыми — гетерохроматиновые. Существует несколько модификаций метода R-окраски и каждый из них имеет свою кодировку по международной цитогенетической номенклатуре. Одним из них является обработка препаратов Ва(ОН)2 с контролируемой тепловой денатурацией и последующей отмывкой в дистиллированной воде и окрашиванием раствором красителя Гимза. Кодировка R-окраски, полученной таким способом — RHG.
С-окраска(от англ. Constitutive heterochromatin — конститутивный гетерохроматин) выявляется в виде вариабельных по величине темноокрашенных сегментов конститутивного гетерохроматина в прицентромерных районах хромосом, в то время как эухроматиновые участки хромосом прокрашиваются очень бледно. Методы получения С-окраски могут варьировать, но важным условием является предварительная обработка препаратов щелочью с последующей двухчасовой инкубацией препарата в двукратном стандартном солевом растворе при 65°С. Окраску препаратов производят красителем Гимза. С-окраска имеет свою кодировку (CBG) по международной цитогенетической номенклатуре. Конститутивный гетерохроматин построен преимущественно из многократно повторяющихся последовательностей ДНК, так называемой сателлитной ДНК, выявляемой во время градиентного центрифугирования хроматина. В популяциях человека, также как и в случае Q-окрашивания, существует межиндивидуальная вариабельность по определенным участкам хромосом, в данном случае по величине блоков С-гетерохроматина, выявляемых с помощью С-окраски (С-полиморфизм хромосом). По локализации выделяют четыре типа С-хроматина: 1) собственно центромерный, присущий всем хромосомам, с относительно небольшими по площади темноокрашенными блоками; 2) более крупные блоки гетерохроматина, располагающиеся в прицентромерных районах длинных плечей аутосом 1, 9 и 16, обладающие четко выраженным межиндивидуальным полиморфизмом; 3) большой блок гетерохроматина дистальной части длинного плеча Y-хромосомы, заметно варьирующий по величине у разных лиц мужского пола; 4) гетерохроматин коротких плеч акроцентрических хромосом. Метод выявления центромерного гетерохроматина позволяет прежде всего оценить хромосомный полиморфизм четырех хромосом набора -1, 9, 16 и Y, а также используется в практике клинической цитогенетики для уточнения характера структурных перестроек, затрагивающих данные хромосомы. По международной цитогенетической номенклатуре увеличенные или уменьшенные блоки гетерохроматина обозначаются как qh+ или qh- (h гетерохроматин). Например, запись кариотипа 46,XX,9qh+ означает, что у нормальной женщины имеется вариант хромосомы 9 с большим гетерохроматиновым блоком в длинном плече, а запись 46,XYqh- означает, что у мужчины имеется уменьшение длины гетерохроматинового блока на длинном плече Y-хромосомы.
NOR-окраска(от англ. Nucleolar Organizer Region — Ядрышко-Образующие Районы — ЯОР) или Ag-окраска (серебрение) применяется для выявления ядрышкообразующих районов, расположенных в коротких плечах (сегмент ql2 — спутничная нить) всех 5 пар акроцентрических хромосом человека (13, 14, 15, 21 и 22), с помощью окрашивания солями серебра. Известно, что районы ядрышковых организаторов содержат гены рибосомальной РНК (рРНК). Единица транскрипции генов рРНК содержит гены 5,8S, 18S и 28S РНК. Ряды рРНК транскрипционных единиц располагаются тандемно вдоль ДНК и отделяются друг от друга нетранскрибируемыми последовательностями — спейсерами. Транскрибируемый район и нетранскрибируемый спейсер тандемно повторяются примерно 40 раз в каждой из 5 пар акроцентрических хромосом, составляя около 400 копий рибосомных генов на геном. В настоящее время для визуализации этих районов на метафазных хромосомах применяют метод серебрения Хоуэлла и Блэка (1980 г.). С помощью этого метода сами хромосомы окрашиваются в желтый цвет, а в коротких плечах акроцентрических хромосом на спутничных нитях четко выделяются черные точечные образования — глыбки восстановленного металлического серебра. Размеры этих глыбок, отражающие активность ЯОР, которые функционировали в предшествующей интерфазе, на разных хромосомах существенно варьируют — от отсутствия заметной окраски — до достаточно крупных блоков серебра (Ag-полиморфизм). Тонкие механизмы окраски ядрышкообразующих районов хромосом пока неизвестны, но ясно, что красящим субстратом является не ДНК рибосомных генов и не рРНК, а кислые белки, связанные с ней. Для оценки данного типа хромосомного полиморфизма предложена полуколичественная 5-балльная система оценки активности ЯОР каждой из пяти пар акроцентрических хромосом генома человека (0 — отсутствие окраски, 1 — слабая, 2 — средняя, 3 — сильная и 4 — очень сильная окраска ЯОР хромосом, при которой размеры Ag-блока значительно превышают толщину хроматид). При сложении баллов отдельных ЯОР может быть оценена активность всех 10 ЯОР генома по критерию суммарной функциональной активности ЯОР, предложенному Н.А. Ляпуновой с соавт. (1988). В популяциях человека существует межиндивидуальный полиморфизм Ag-окрашенных хромосом, который выражается в специфичности степени окраски отдельных хромосом у разных индивидов, являющейся наследуемой характеристикой.
Т-окраска(от англ. Telomere — теломера) — применяется для выявления теломерных районов хромосом в коротких и длинных плечах. Метод включает инкубацию препаратов хромосом при 87°С в течение 20-60 мин в растворе фосфатного буфера при рН 5,1 с последующей окраской раствором красителя Гимза.
Источник



