
- полуконсервативная репликация
- Смотреть что такое «полуконсервативная репликация» в других словарях:
- Полуконсервативный способ репликации
- Полуконсервативный способ репликации ДНК. Биологическое значение.
- Полуконсервативная репликация — Semiconservative replication
- СОДЕРЖАНИЕ
- Открытие
- Модели репликации
- Разделение и рекомбинация двухцепочечной ДНК
- Скорость и точность
- Приложения
полуконсервативная репликация
Смотреть что такое «полуконсервативная репликация» в других словарях:
полуконсервативная репликация — Метод репликации молекулы ДНК, при которой каждая материнская цепь молекулы ДНК достраивает себе новую дочернюю цепь [http://www.dunwoodypress.com/148/PDF/Biotech Eng Rus.pdf] Тематики биотехнологии EN semiconservative replication … Справочник технического переводчика
Полуконсервативная репликация — * паўкансерватыўная рэплікацыя * semiconservative replication тип репликации ДНК, при котором молекула делится продольно, каждая половина сохраняется и служит матрицей для образующейся новой нити. Термин был введен, когда точный процесс… … Генетика. Энциклопедический словарь
Репликация полуконсервативная — * рэплікацыя паўкансерватыўная * semi conservative replication процесс репликации, при котором дочерние клетки первого поколения получают одну цепь ДНК от родителей, а вторая цепь синтезируется вновь. Такой же процесс повторяется при образовании… … Генетика. Энциклопедический словарь
репликация θ-типа — репликация θ типа θ репликация Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования «вздутия», видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Справочник технического переводчика
θ-репликация — θ type replication репликация θ типа, θ репликация. Двунаправленная полуконсервативная репликация кольцевых молекул ДНК, начинающаяся с образования вздутия , видимого под электронным микроскопом, расширяющегося в двух направлениях; перед… … Молекулярная биология и генетика. Толковый словарь.
Семиконсервативная репликация — * семікансерватыўная рэплікацыя * semiconservative replication … Генетика. Энциклопедический словарь
генетический код — Долгое время наследственные механизмы были предметом внимания генетиков, но природа молекул, переносящих информацию от одного индивида к другому, оставалась неведомой. Была известна роль макромолекул протеина и нуклеиновых кислот в этом… … Западная философия от истоков до наших дней
История науки — История науки это исследование феномена науки в его истории. Наука, в частности, представляет собой совокупность эмпирических, теоретических и практических знаний о Мире, полученных научным сообществом. Поскольку с одной стороны наука… … Википедия
semiconservative replication — semiconservative replication. См. полуконсервативная репликация. (Источник: «Англо русский толковый словарь генетических терминов». Арефьев В.А., Лисовенко Л.А., Москва: Изд во ВНИРО, 1995 г.) … Молекулярная биология и генетика. Толковый словарь.
5 -гидроксиметилцитозин 5 -гМц — 5 гидроксиметилцитозин, 5 гМц * 5 гідроксіметылцытазін * 5 hydroxymethyl cytosine or 5 hMC пиримидиновое основание, замещающее цитозин в ДНК четных колифагов. 5 ГМЦ комплементарен гуанину. Считается, что фагоспецифичная ДНКаза бактериофага… … Генетика. Энциклопедический словарь
Источник
Полуконсервативный способ репликации




![]()
![]()
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.
Гипотеза Уотсона-Крика была проверена с помощью опытов, выполненных М.Мезельсоном и Ф.Сталем в 1957 г. Клетки E.coli выращивали в течение ряда поколений в среде, содержащей в качестве источника азота хлористый аммоний NH4Cl, в котором обычный изотоп [ 14 N] был заменен на «тяжелый» изотоп [ 15 N]. Вследствие этого все соединения клеток, имеющие в своем составе азот, в том числе и азотистые основания ДНК, оказались обогащенными изотопом [ 15 N]. Плотность ДНК, выделенной из этих клеток, была выше плотности нормальной [ 14 N] ДНК. Смесь «тяжелой» [ 15 N] и «легкой» [ 14 N] ДНК удалось разделить методом центрифугирования в концентрированном растворе хлористого цезия. Поскольку [ 15 N] ДНК чуть тяжелее, чем [ 14 N] ДНК, полоса, в которой она достигает равновесия в градиенте СsCl, расположена ближе ко дну пробирки, чем полоса с [ 14 N] ДНК (рис.7)
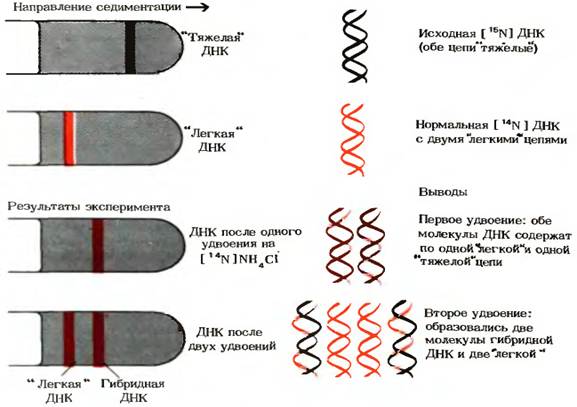
Рис. 7 Результаты эксперимента Мезельсона-Сталя
Мезельсон и Сталь перенесли клетки E.coli, росшие на среде с изотопом [ 15 N] и содержащие «тяжелые» цепи ДНК, на свежую среду с обычным изотопом [ 14 N]. На этой среде клетки E.coli выращивали в течение времени, необходимого для удвоения клеток. Затем из этих клеток выделяли ДНК и анализировали ее плотность с помощью описанного выше метода седиментации (осаждения ДНК в растворе СsCl). В градиенте СsCl была обнаружена лишь одна полоса ДНК, плотность которой оказалась средней между плотностью нормальной «легкой» [ 14 N] ДНК и плотностью «тяжелой» [ 15 N] ДНК (рис. ). Это подтвердило, что двухцепочечная ДНК дочерних клеток содержит одну старую 15 N – цепь от родительской ДНК и одну новую 14 N – цепь.
Если выделить ДНК из клеток, которые прошли два цикла удвоения на среде с [ 14 N], то она разделится на две полосы: одна с плотностью, соответствующей плотности нормальной «легкой» ДНК, а другая с плотностью «гибридной» ДНК, наблюдавшейся после первого удвоения клеток. На основании этих данных Мезельсон и Сталь пришли к выводу, что в строгом соответствии с гипотезой Уотсона-Крика каждый дочерний дуплекс ДНК после двух циклов удвоения клеток содержал одну родительскую и одну новообразованную цепь ДНК. Такой механизм репликации назвали полуконсервативным. Полученные результаты полностью исключили консервативный способ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсон и Сталя позволил также отвергнуть дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из коротких участков как родительской, так и новообразованнй ДНК, соединенных между собой случайным образом.
Источник
Полуконсервативный способ репликации ДНК. Биологическое значение.
Полуконсервативный механизм репликации ДНК. Перед каждым делением клетки в ней должно удвоиться содержание ДНК, чтобы каждая дочерняя клетка получила полный набор хромосом. Основу каждой хромосомы образует одна двухцепочечная молекула ДНК. Предложенная Дж. Уотсоном и Ф. Криком модель строения ДНК форме регулярной двойной спирали сразу же позволила понять принцип копирования ДНК. Ее репликация происходит полуконсервативным способом: две исходные цепи материнской ДНК расходятся, и каждая из них становится матрицей для синтеза новой комплементарной цепи. Таким образом, каждая новая двойная спираль ДНК содержит одну старую и одну новую цепь. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух материнский цепей ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсон и Ф. Сталь.
Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся и каждая цепь служит матрицей для синтеза новой комплементарной цепи ДНК. Каждая дочерняя молекула состоит из одной старой материнской и одной новой синтезированной цепи ДНК.
Общие принципы репликации ДНК.
В основе процесса репликации лежит принцип копирования материнской цепи ДНК с образованием двух идентичных молекул ДНК. В основе синтеза новой цепи ДНК лежит принцип комплементарности азотистых оснований, т.е. последовательность нуклеотидов материнской цепи определяет последовательность нуклеотидов в синтезируемой цепи ДНК
Синтез новых цепей ДНК идет в направлении 5’ → 3’.
В основе репликации ДНК лежат принципы антипараллельности и униполярности. Синтез новых цепей ДНК идет в направлении от 5’-конца к 3’ — концу, при этом ДНК-полимеразы – ферменты, синтезирующие новые цепи ДНК, — добавляют нуклеотиды к 3’- концу наращиваемой цепи ДНК. При этом матричная цепь имеет противоположную ориентацию: 3’ → 5’, т.е. ДНК-полимеразы могут передвигаться вдоль матрицы только в направлении от 3’ к 5’-концам.
Механизмы синтеза ДНК хорошо изучены в клетках бактерий, однако имеются доказательства, что в клетках эукариот процесс протекает аналогичным образом.
Инициация репликации ДНК. Репликация начинается в специфическом участке молекулы ДНК, который называется точка начала репликации или ориджин.
Точка начала репликации (origin) – это участок молекулы ДНК со специфической последовательностью нуклеотидов с большим содержанием пар АТ (последовательность 300 п.н., богата АТ). Специальные инициирующие белки необходимы для связывания ферментов репликации с молекулой ДНК: белок DnaA – для прокариот белок RPA (replication protein A) – для эукриот.
Кольцевая хромосома прокариот имеет одну точку начала репликации, которая
называется OriC. В этой точке цепи расходятся и образуются две репликативные вилки, которые движутся в противоположном направлении. Скорость синтеза ДНК в клетках прокариот составляет 500 нукл./сек. Две вилки встречаются на противоположной стороне кольца. В клетках прокариот существует специальный фермент гираза (топоизомераза II), который разделяет две образующиеся кольцевые молекулы ДНК. Антибиотик налидиксовая кислота угнетает размножение бактерий, путем инактивации гиразы.
В клетках эукариот этот фермент отсутствует, поэтому налидиксовую кислоту используют в клинической практике для лечения бактериальных инфекций.
Репликация ДНК эукариот начинается одновременно во многих точках начала репликации, от каждой точки движутся две репликативные вилки в противоположных направлениях. Скорость синтеза молекул ДНК эукариот составляет 50 нукл./сек.
Репликон – фрагмент молекулы ДНК, репликация которого происходит под контролем одной точки начала репликации. Кольцевая хромосома прокариот имеет 1 репликон. Геномы эукариот содержат сотни и тысячи репликонов.
Инициация — образование репликативной вилки. Нити ДНК разделяются благодаря действию специальных ферментов и белков.
Хеликаза (от англ. helix – спираль) — основной фермент, расплетающий цепи ДНК. У прокариот он называется белок DnaB. Хеликаза разрывает водородные связи между комплементарными основаниями, используя энергию АТФ.
Топоизомеразы – ферменты, которые устраняют положительные сверхвитки перед репликативной вилкой. Эти ферменты временно разрывают нити материнской ДНК в двойной спирали перед репликативной вилкой, после завершения процесса нити ДНК восстанавливают целостность.
SSB — белки – это белки, которые связываются с одноцепочечной ДНК и удерживают матрицу. В результате образуется репликативная вилка, где и происходит синтез новых цепей ДНК.
Источник
Полуконсервативная репликация — Semiconservative replication
Полуконсервативная репликация описывает механизм репликации ДНК во всех известных клетках. Репликация ДНК происходит в нескольких точках репликации вдоль цепи матрицы ДНК. Поскольку двойная спираль ДНК разматывается геликазой , репликация происходит отдельно на каждой цепи матрицы в антипараллельных направлениях. Этот процесс известен как полуконсервативная репликация, потому что создаются две копии исходной молекулы ДНК. Каждая копия содержит одну исходную цепочку и одну вновь синтезированную цепочку. Структура ДНК (расшифрованная Джеймсом Д. Уотсоном и Фрэнсисом Криком в 1953 г.) предполагала, что каждая цепь двойной спирали будет служить шаблоном для синтеза новой цепи. Неизвестно, как вновь синтезированные цепи объединяются с цепями-матрицами с образованием двух двойных спиральных молекул ДНК.
СОДЕРЖАНИЕ
Открытие
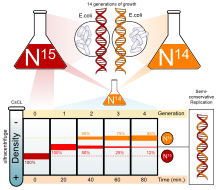
Было проведено множество экспериментов, чтобы определить, как реплицируется ДНК. Полуконсервативная модель была предвосхищена Николаем Кольцовым и позже подтверждена экспериментом Мезельсона-Шталя ., Который подтвердил, что ДНК реплицируется полуконсервативно путем проведения эксперимента с использованием двух изотопов : азота-15 ( 15
N
) и азот-14 ( 14
N
). Когда 14
N
был добавлен к тяжелому 15
N
— 15
N
ДНК, гибрид 15
N
— 14
N
был замечен в первом поколении. После второго поколения гибрид остался, но легкая ДНК ( 14
N
— 14
N
) тоже было замечено. Это указывало на полуконсервативную репликацию ДНК. Этот режим репликации ДНК позволял каждой дочерней цепи оставаться связанной со своей цепочкой-матрицей.
Модели репликации
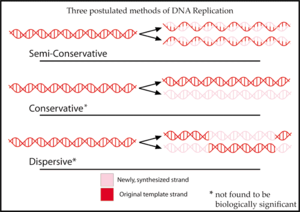
Полуконсервативная репликация получила свое название от того факта, что этот механизм транскрипции был одной из трех моделей, первоначально предложенных для репликации ДНК :
- Полуконсервативная репликация даст две копии, каждая из которых содержит одну из исходных цепей ДНК и одну новую цепочку. Полуконсервативная репликация полезна для восстановления ДНК. Во время репликации новая цепь ДНК приспосабливается к модификациям, внесенным в цепочку-матрицу.
- Консервативная репликация оставит две исходные цепи ДНК-матрицы вместе в двойной спирали и создаст копию, состоящую из двух новых цепей, содержащих все новые пары оснований ДНК.
- Дисперсионная репликация создаст две копии ДНК, каждая из которых содержит отдельные участки ДНК, состоящие либо из обеих исходных цепей, либо из обеих новых цепей. Первоначально считалось, что цепи ДНК разрываются на каждой десятой паре оснований, чтобы добавить новую матрицу ДНК. В конце концов, вся новая ДНК будет образовывать двойную спираль после многих поколений репликации.
Разделение и рекомбинация двухцепочечной ДНК
Для полуконсервативной репликации двойную спираль ДНК необходимо разделить, чтобы новая цепь матрицы могла быть связана с комплементарными парами оснований. Топоизомераза — это фермент, который способствует распаковке и рекомбинации двойной спирали. В частности, топоизомераза предотвращает сверхспирализацию двойной спирали или ее слишком тугое наматывание. Три топоизомеразы ферменты участвуют в этом процессе: Тип IA топоизомеразы , тип IB топоизомеразы и Type II топоизомеразы . Топоизомераза типа I раскручивает двухцепочечную ДНК, в то время как топоизомераза типа II разрывает водородные связи, связывающие комплементарные пары оснований ДНК.
Скорость и точность
Скорость полуконсервативной репликации ДНК в живой клетке сначала измеряли как скорость удлинения цепи ДНК фага Т4 в инфицированной фагом E. coli . В период экспоненциального увеличения ДНК при 37 ° C скорость удлинения цепи составляла 749 нуклеотидов в секунду. Частота мутаций на пару оснований за раунд репликации во время синтеза ДНК фага Т4 составляет 2,4 × 10 −8 . Таким образом, полуконсервативная репликация ДНК происходит быстро и точно.
Приложения
Полуконсервативная репликация дает ДНК множество преимуществ. Это быстро, точно и позволяет легко восстанавливать ДНК. Он также отвечает за фенотипическое разнообразие у нескольких видов прокариот. Процесс создания вновь синтезированной цепи из цепи-шаблона позволяет метилировать старую цепь в отдельное время от новой цепи. Это позволяет репаративным ферментам проверять новую цепь и исправлять любые мутации или ошибки.
ДНК может иметь способность активировать или деактивировать определенные области на вновь синтезированной цепи, что позволяет изменять фенотип клетки. Это может быть выгодно для клетки, потому что ДНК может активировать более благоприятный фенотип, чтобы помочь в выживании. Благодаря естественному отбору более благоприятный фенотип будет сохраняться у всего вида. Это порождает идею наследования или того, почему одни фенотипы наследуются над другими.
Источник



