
Полуконсервативный способ репликации




![]()
![]()
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.
Гипотеза Уотсона-Крика была проверена с помощью опытов, выполненных М.Мезельсоном и Ф.Сталем в 1957 г. Клетки E.coli выращивали в течение ряда поколений в среде, содержащей в качестве источника азота хлористый аммоний NH4Cl, в котором обычный изотоп [ 14 N] был заменен на «тяжелый» изотоп [ 15 N]. Вследствие этого все соединения клеток, имеющие в своем составе азот, в том числе и азотистые основания ДНК, оказались обогащенными изотопом [ 15 N]. Плотность ДНК, выделенной из этих клеток, была выше плотности нормальной [ 14 N] ДНК. Смесь «тяжелой» [ 15 N] и «легкой» [ 14 N] ДНК удалось разделить методом центрифугирования в концентрированном растворе хлористого цезия. Поскольку [ 15 N] ДНК чуть тяжелее, чем [ 14 N] ДНК, полоса, в которой она достигает равновесия в градиенте СsCl, расположена ближе ко дну пробирки, чем полоса с [ 14 N] ДНК (рис.7)
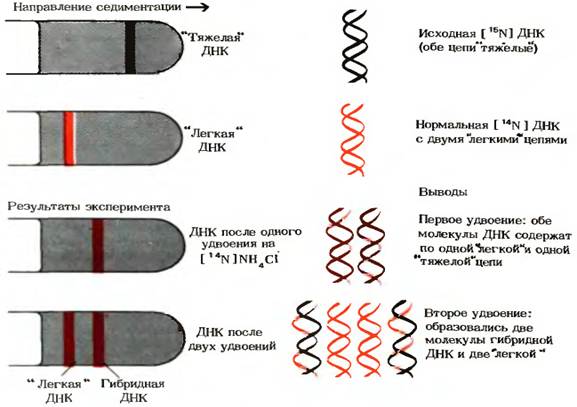
Рис. 7 Результаты эксперимента Мезельсона-Сталя
Мезельсон и Сталь перенесли клетки E.coli, росшие на среде с изотопом [ 15 N] и содержащие «тяжелые» цепи ДНК, на свежую среду с обычным изотопом [ 14 N]. На этой среде клетки E.coli выращивали в течение времени, необходимого для удвоения клеток. Затем из этих клеток выделяли ДНК и анализировали ее плотность с помощью описанного выше метода седиментации (осаждения ДНК в растворе СsCl). В градиенте СsCl была обнаружена лишь одна полоса ДНК, плотность которой оказалась средней между плотностью нормальной «легкой» [ 14 N] ДНК и плотностью «тяжелой» [ 15 N] ДНК (рис. ). Это подтвердило, что двухцепочечная ДНК дочерних клеток содержит одну старую 15 N – цепь от родительской ДНК и одну новую 14 N – цепь.
Если выделить ДНК из клеток, которые прошли два цикла удвоения на среде с [ 14 N], то она разделится на две полосы: одна с плотностью, соответствующей плотности нормальной «легкой» ДНК, а другая с плотностью «гибридной» ДНК, наблюдавшейся после первого удвоения клеток. На основании этих данных Мезельсон и Сталь пришли к выводу, что в строгом соответствии с гипотезой Уотсона-Крика каждый дочерний дуплекс ДНК после двух циклов удвоения клеток содержал одну родительскую и одну новообразованную цепь ДНК. Такой механизм репликации назвали полуконсервативным. Полученные результаты полностью исключили консервативный способ репликации, при котором одна дочерняя ДНК должна была бы содержать обе исходные цепи, а другая состояла бы из двух новосинтезированных цепей. Опыт Мезельсон и Сталя позволил также отвергнуть дисперсивный механизм репликации, при котором каждая дочерняя цепь ДНК состоит из коротких участков как родительской, так и новообразованнй ДНК, соединенных между собой случайным образом.
Источник
Биология. 11 класс
§ 16. Клеточный цикл. Репликация ДНК
Клеточный цикл . Все новые клетки образуются путем деления уже существующих. Этот принцип, как уже отмечалось, сформулировал Р. Вирхов еще в середине XIX в. Деление клеток обеспечивает непрерывность существования жизни на нашей планете. Именно благодаря ему осуществляются различные способы бесполого и полового размножения организмов. В основе процессов роста, регенерации и индивидуального развития многоклеточных организмов также лежит деление клеток.
Период существования клетки от момента ее образования из материнской клетки до собственного деления (включая это деление) или гибели называется клеточным циклом.
Продолжительность клеточного цикла у разных организмов и различных клеток в составе одного организма варьирует. Так, у бактерий в благоприятных условиях он длится примерно 20 мин. Короткие клеточные циклы (30—60 мин) характерны для бластомеров рыб и земноводных на этапе дробления, в то время как у млекопитающих промежуток времени между делениями бластомеров может достигать 10 ч и более. У взрослых мышей клетки кишечного эпителия делятся каждые 11—22 ч, а роговицы глаза — приблизительно один раз в трое суток. Для регулярно делящихся клеток многоклеточных организмов длительность клеточного цикла обычно составляет 12—36 ч.
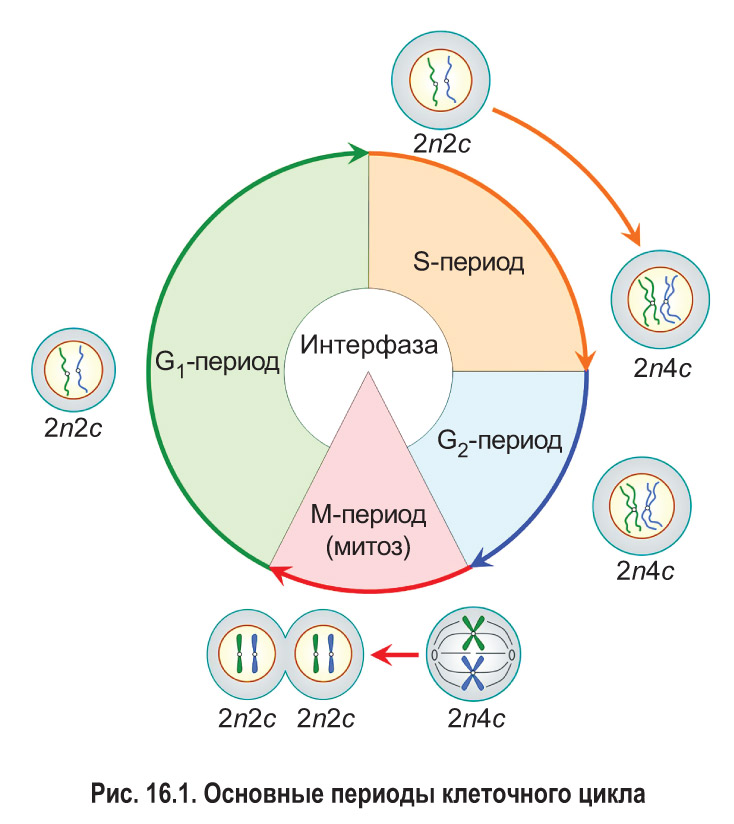 Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Клеточный цикл состоит из интерфазы и деления клетки (рис. 16.1). Интерфаза — это часть клеточного цикла между двумя последовательными делениями. Как правило, она занимает больше времени, чем само деление. Рассмотрим основные периоды интерфазы на примере эукариотической клетки.
Пресинтетический, или G1-пе риод (от англ. gap — промежуток), начинается с момента образования новой клетки в результате деления материнской. Обычно это самый длительный период интерфазы и клеточного цикла в целом. В течение G1-периода молодая клетка интенсивно растет, в ней увеличивается количество органоидов и синтезируются различные соединения, необходимые для протекания процессов жизнедеятельности. В том числе образуются вещества, которые будут нужны для последующего удвоения молекул ДНК.
Вы уже знаете, что набор хромосом обозначают как n: например, 1n — гаплоидный набор, 2n — диплоидный. Набор молекул ДНК в клетках принято записывать с помощью буквы с. Из § 14 вам известно, что каждая хроматида содержит одну молекулу ДНК, т. е. количество молекул ДНК и хроматид в составе хромосом всегда совпадает. Таким образом, записи типа 1с, 2с, 4с отражают содержание в клетках не только молекул ДНК, но и соответствующих хроматид.
В пресинтетическом периоде каждая хромосома состоит из одной хроматиды. Следовательно, в G1-периоде число хромосом (n) и хроматид (с) в клетке одинаковое. Набор хромосом и хроматид диплоидной клетки в G1-периоде клеточного цикла можно выразить записью 2n2c.
В синтетическом, или S-периоде (от англ. synthesis — синтез), происходит удвоение молекул ДНК — репликация, а также удвоение центриолей клеточного центра (в тех клетках, где он имеется). После завершения репликации каждая хромосома состоит уже из двух идентичных сестринских хроматид, которые соединены друг с другом в области центромеры. Количество хроматид в каждой паре гомологичных хромосом становится равным четырем. Таким образом, набор хромосом и хроматид диплоидной клетки в конце S-периода (т. е. после репликации) выражается записью 2n4c.
Постсинтетический, или G2-период, продолжается от окончания синтеза ДНК (репликации) до начала деления клетки. В это время клетка активно запасает энергию и синтезирует белки для предстоящего деления (например, белок тубулин для построения микротрубочек, образующих впоследствии веретено деления). В течение всего G2-периода набор хромосом и хроматид в клетке остается неизменным — 2n4c.
Итак, интерфаза обычно включает три периода: пресинтетический (G1), синтетический (S) и постсинтетический (G2). На протяжении всей интерфазы хромосомы не спирализованы. Они располагаются в ядре клетки в виде хроматина .
После завершения интерфазы начинается деление клетки. Основным способом деления клеток эукариот является митоз, поэтому данный этап клеточного цикла обозначают как М-период. При митозе происходит спирализация хроматина . Это приводит к формированию компактных двухроматидных хромосом. После этого сестринские хроматиды каждой хромосомы отделяются друг от друга и затем попадают в разные дочерние клетки. Значит, дочерние клетки, образовавшиеся в результате митоза и вступающие в новый клеточный цикл, имеют набор 2n2c.
Обобщенная информация об основных периодах клеточного цикла представлена в таблице 16.1.
Таблица 16.1. Основные периоды клеточного цикла
Период
Содержание наследственного материала в диплоидной клетке
Источник
Полуконсервативный способ репликации ДНК. Биологическое значение.
Полуконсервативный механизм репликации ДНК. Перед каждым делением клетки в ней должно удвоиться содержание ДНК, чтобы каждая дочерняя клетка получила полный набор хромосом. Основу каждой хромосомы образует одна двухцепочечная молекула ДНК. Предложенная Дж. Уотсоном и Ф. Криком модель строения ДНК форме регулярной двойной спирали сразу же позволила понять принцип копирования ДНК. Ее репликация происходит полуконсервативным способом: две исходные цепи материнской ДНК расходятся, и каждая из них становится матрицей для синтеза новой комплементарной цепи. Таким образом, каждая новая двойная спираль ДНК содержит одну старую и одну новую цепь. Такой механизм репликации ДНК, при котором от одного поколения к другому передается одна из двух материнский цепей ДНК, получил название полуконсервативного и был экспериментально доказан в 1958 году М. Мезельсон и Ф. Сталь.
Легко представить, что удвоение ДНК происходит вследствие того, что цепи расходятся и каждая цепь служит матрицей для синтеза новой комплементарной цепи ДНК. Каждая дочерняя молекула состоит из одной старой материнской и одной новой синтезированной цепи ДНК.
Общие принципы репликации ДНК.
В основе процесса репликации лежит принцип копирования материнской цепи ДНК с образованием двух идентичных молекул ДНК. В основе синтеза новой цепи ДНК лежит принцип комплементарности азотистых оснований, т.е. последовательность нуклеотидов материнской цепи определяет последовательность нуклеотидов в синтезируемой цепи ДНК
Синтез новых цепей ДНК идет в направлении 5’ → 3’.
В основе репликации ДНК лежат принципы антипараллельности и униполярности. Синтез новых цепей ДНК идет в направлении от 5’-конца к 3’ — концу, при этом ДНК-полимеразы – ферменты, синтезирующие новые цепи ДНК, — добавляют нуклеотиды к 3’- концу наращиваемой цепи ДНК. При этом матричная цепь имеет противоположную ориентацию: 3’ → 5’, т.е. ДНК-полимеразы могут передвигаться вдоль матрицы только в направлении от 3’ к 5’-концам.
Механизмы синтеза ДНК хорошо изучены в клетках бактерий, однако имеются доказательства, что в клетках эукариот процесс протекает аналогичным образом.
Инициация репликации ДНК. Репликация начинается в специфическом участке молекулы ДНК, который называется точка начала репликации или ориджин.
Точка начала репликации (origin) – это участок молекулы ДНК со специфической последовательностью нуклеотидов с большим содержанием пар АТ (последовательность 300 п.н., богата АТ). Специальные инициирующие белки необходимы для связывания ферментов репликации с молекулой ДНК: белок DnaA – для прокариот белок RPA (replication protein A) – для эукриот.
Кольцевая хромосома прокариот имеет одну точку начала репликации, которая
называется OriC. В этой точке цепи расходятся и образуются две репликативные вилки, которые движутся в противоположном направлении. Скорость синтеза ДНК в клетках прокариот составляет 500 нукл./сек. Две вилки встречаются на противоположной стороне кольца. В клетках прокариот существует специальный фермент гираза (топоизомераза II), который разделяет две образующиеся кольцевые молекулы ДНК. Антибиотик налидиксовая кислота угнетает размножение бактерий, путем инактивации гиразы.
В клетках эукариот этот фермент отсутствует, поэтому налидиксовую кислоту используют в клинической практике для лечения бактериальных инфекций.
Репликация ДНК эукариот начинается одновременно во многих точках начала репликации, от каждой точки движутся две репликативные вилки в противоположных направлениях. Скорость синтеза молекул ДНК эукариот составляет 50 нукл./сек.
Репликон – фрагмент молекулы ДНК, репликация которого происходит под контролем одной точки начала репликации. Кольцевая хромосома прокариот имеет 1 репликон. Геномы эукариот содержат сотни и тысячи репликонов.
Инициация — образование репликативной вилки. Нити ДНК разделяются благодаря действию специальных ферментов и белков.
Хеликаза (от англ. helix – спираль) — основной фермент, расплетающий цепи ДНК. У прокариот он называется белок DnaB. Хеликаза разрывает водородные связи между комплементарными основаниями, используя энергию АТФ.
Топоизомеразы – ферменты, которые устраняют положительные сверхвитки перед репликативной вилкой. Эти ферменты временно разрывают нити материнской ДНК в двойной спирали перед репликативной вилкой, после завершения процесса нити ДНК восстанавливают целостность.
SSB — белки – это белки, которые связываются с одноцепочечной ДНК и удерживают матрицу. В результате образуется репликативная вилка, где и происходит синтез новых цепей ДНК.
Источник
Вопрос 19. Репликация ДНК (полуконсервативный механизм)
Синтез ДНК в подавляющем большинстве случаев проходит по полуконсервативному механизму , то есть дочерняя цепь ДНК синтезируется по материнской цепи, называемой обычно матрицей. В одних случаях матрицами являются односпиральные ДНК, в других — РНК.
Репликация, редупликация, ауторепродукция, протекающий во всех живых клетках процесс самовоспроизведения нуклеиновых кислот, генов, хромосом.
Репликация ДНК — это процесс синтеза дочерней молекулы дезоксирибонуклеиновой кислоты, который происходит в процессе деления клетки на матрице родительской молекулы ДНК. При этом генетический материал, зашифрованный в ДНК, удваивается и делится между дочерними клетками. Репликацию ДНК осуществляет фермент ДНК-полимераза.
В основе механизма репликация лежит ферментативный синтез дезоксирибонуклеиновой кислоты (ДНК) или рибонуклеиновых кислот (РНК), осуществляемый по матричному принципу. Строгая специфичность спаривания азотистых оснований в молекуле ДНК обусловливает комплементарность последовательностей оснований в двух цепях и обеспечивает высокую точность Репликация Пары гуанин — цитозин стабилизируется тремя водородными связями, пара аденин — тимин — двумя. Согласно Уотсону и Крику, процесс Репликация ДНК предусматривает: 1) разрыв водородных связей и расплетение нитей двойной спирали; 2) синтез на одиночных нитях комплементарных цепей. В результате из одной двухцепочечной ДНК возникают две подобные молекулы, причём в каждой из дочерних молекул одна полинуклеотидная цепь родительская, а другая — синтезированная заново (полуконсервативный механизм Репликация).
У вирусов и фагов, имеющих однонитевую ДНК, Репликация идёт особым образом. После внедрения в клетку хозяина одноцепочечной ДНК, которую называют (+)-цепью, на ней, как на матрице, синтезируется комплементарная ей (—)-цепь. На образовавшейся двухспиральной молекуле (репликативная форма) синтезируются новые одноцепочечные (+)-цепи, включающиеся в новые вирусные частицы. По такому же принципу происходит Репликация РНК-содержащих вирусов и фагов.
У высших организмов — эукариотов, клетки которых содержат сформированное ядро, основную генетическую функцию несут сложно организованные структуры — хромосомы, состоящие из ДНК, РНК, белков и других веществ. В интерфазе, предшествующей делению клеток, осуществляется Репликация ДНК и других компонентов хромосом; затем удвоенные хромосомы разъединяются и распределяются равномерно между дочерними клетками. Т. о., вся наследственная информация в относительно неизмененном виде передаётся от клетки к клетке, от поколения к поколению.
Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Репликация у эукариот осуществляется несколькими разными ДНК-полимеразами. Далее происходит закручивание синтезированных молекул по принципу суперспирализации и дальнейшей компактизации ДНК. Синтез энергозатратный.
Цепи молекулы ДНК расходятся, образую репликационную вилку, и каждая из них становится матрицей, на которой синтезируется новая комплементарная цепь. В результате образуются две новые двуспиральные молекулы ДНК, идентичные родительской молекуле.
Каждая молекула ДНК состоит из одной цепи исходной родительской молекулы и одной вновь синтезированной цепи. Такой механизм репликации называется полуконсервативным.
Процесс репликации ДНК
Репликация состоит из большого числа последовательных этапов, которые включают узнавание точки началу репликации, расплетание исходного дуплекса (спирали), удержание его цепей в изолированном друг от друга состоянии, инициацию синтеза на них новых дочерних цепей, их рост (элонгацию), закручивание цепей в спираль и терминацию (окончание) синтеза. Функциональная единица репликации – репликон, представляющий собой сегмент (участок) хромосомы или внехромосомной ДНК, ограниченный точкой начала, в которой инициируется репликация, и точкой окончания, в которой репликация останавливается. Скорость репликации контролируется на стадии инициации. Однажды начавшись, репликация продолжается до тех пор, пока весь репликон не будет дуплицирован (удвоен). В каждом клеточном цикле репликация инициируется только один раз. Начиная с точки инициации, репликация осуществляется в ограниченной зоне, перемещающейся вдоль исходной спирали ДНК. Эта активная зона репликации (т.н. репликац. вилка) может двигаться в обоих направлениях. При однонаправленной репликации вдоль ДНК движется одна репликационная вилка. При двунаправленной репликации от точки инициации в противоположных направлениях расходятся две репликационные вилки; Движение двух вилок в противоположных направлениях создает петлю, которая имеет вид «пузыря» или «глаза». Продолжающаяся репликация расширяет «глаз» до тех пор, пока он не включит в себя весь репликон.
В ходе репликации рост цепи осуществляется благодаря взаимодействию дезоксирибонуклеозидтрифосфата с 3′-ОН концевым нуклеотидом уже построенной части ДНК; при этом отщепляется пирофосфат и образуется фосфодиэфирная связь. Рост полинуклеотидной цепи идет только с ее З’-конца, т. е. в направлении 5′ : 3′. Фермент, катализирующий эту реакцию, -ДНК – полимераза.
ДНК-полимераза имеет один центр связывания нуклеозидтрифосфата, общий для всех четырех нуклеотидов.
Репликационная вилка асимметрична. Из двух синтезируемых дочерних цепей ДНК одна строится непрерывно, а другая – с перерывами. Первую называют ведущей, или лидирующей, цепью, а вторую – отстающей. Синтез второй цепи идет медленнее; Благодаря такому прерывистому механизму синтеза, репликация обеих антипараллельных цепей осуществляется с участием одного фермента-ДНК-полимеразы, катализирующего наращивание нуклеотидной цепи только в направлении 5′ : 3′.
В качестве затравок для синтеза фрагментов отстающей цепи служат короткие отрезки РНК, комплементарные матричной цепи ДНК. Эти РНК-затравки (праймеры), состоящие примерно из 10 нуклеотидов, с определенными интервалами синтезируются на матрице отстающей цепи из рибонуклеозидтрифосфатов в направлении 5′ : 3′ с помощью фермента РНК-праймазы. РНК-праймеры затем наращиваются дезоксинуклеотидами с 3′-конца ДНК-полимеразой, которая продолжает наращивание до тех пор, пока строящаяся цепь не достигает РНК-затравки, присоединенной к 5′-концу предыдущего фрагмента.
Завершает весь процесс фермент ДНК-лигаза, катализирующий образование фосфодиэфирной связи между группой З’-ОН нового фрагмента ДНК и 5′-фосфатной группой предыдущего фрагмента. Образование этой связи требует затраты энергии, к-рая поставляется в ходе сопряженного гидролиза пирофосфатной связи кофермента-никотинамид-адениндинуклеотида (в бактериальных клетках) или АТФ (в животных клетках и у бактериофагов).
Геликазы расплетают короткие участки ДНК, находящиеся непосредственно перед репликационной вилкой. На разделение каждой пары оснований расходуется энергия гидролиза двух молекул АТФ до аденозиндифосфата и фосфата. К каждой из разделившихся цепей присоединяется несколько молекул ДНК-связывающих белков, которые препятствуют образованию комплементарных пар и обратному воссоединению цепей.
В случае кольцевого репликона (напр., у плазмиды) описанный процесс наз. q-репликацией. Кольцевые молекулы ДНК закручены сами на себя (суперспирализованы), при раскручивании двойной спирали в процессе репликации они должны непрерывно вращаться вокруг собственной оси. При этом возникает торсионное напряжение, которое устраняется путем разрыва одной из цепей. Затем оба конца сразу же вновь соединяются друг с другом. Эту функцию выполняет фермент ДНК-топоизомераза. Репликация в этом случае обычно происходит в двух направлениях, т.е. существуют две репликационные вилки.
Альтернативный вариант репликации кольцевого репликона предполагает разрыв в одной из цепей двухспиральной молекулы ДНК. Образовавшийся при этом свободный 3′-конец ковалентно наращивается, оставаясь связанным с матрицей (второй, неразорванной цепью), а 5′-конец постепенно вытесняется новой полинуклеотидной цепью. Таким образом одна цепь разматывается и непрерывно удлиняется, а репликационная вилка скользит вокруг кольцевой матричной цепи (механизм «катящегося кольца»). По мере роста новой цепи вытесненная цепь с освободившимся 5′-концом становится линейной матрицей для синтеза новой комплементарной цепи.
Известно несколько типов репликации, РНК:
1. вирусы, содержащие матричные РНК, или мРНК [т. наз. (+)РНК], в результате репликации образуют комплементарную ей цепь [(-)РНК], не являющуюся мРНК, которая используется как матрица для синтеза (+)РНК;
2. вирусы, содержащие (—)РНК, в результате репликации синтезируют (+)РНК;
3. вирусы, содержащие двухцепочечную РНК [(+)PHK и (—)РНК], в результате асимметрической репликации синтезируют (+)РНК.
Гипотеза о механизме репликации сформулирована в 1953 Дж. Уотсоном и Ф. Криком, которые предположили, что две комплементарные цепи ДНК после их разделения могут выполнять функции матриц для образования на них новых цепей ДНК.
Источник



