
Бактериальная конъюгация это способ
Длительное время считали, что бактерии — изолированные генетические системы, и каждая особь имеет одного (и только одного) родителя, то есть их изменчивость вызвана лишь мутациями. Никто не мог себе представить, что, подобно гаметам высших организмов, бактерии способны обмениваться генетическим материалом и, по аналогии с половым размножением, давать начало потомству с новыми свойствами. Однако Лёдерберг и Тёйтем (1946), высевая на минеральную среду, предварительно смешав два типа мутантов Escherichia coli (один нуждался в биотине и метионине, другой — в треонине и лейцине), показали, что у бактерий генетические рекомбинанты со способностью к синтезу всех четырёх факторов роста возникают с частотой 10″6, хотя теоретическая вероятность реверсии по двум генам составляет 10-14-10-16 на генерацию. Это противоречие было снято установлением факта прямой передачи генетического материала от донорной клетки к реципиентной (конъюгация). Помимо конъюгации, передача генетического материала у бактерий может осуществляться также с помощью трансформации и трансдукции (рис. 4-16).
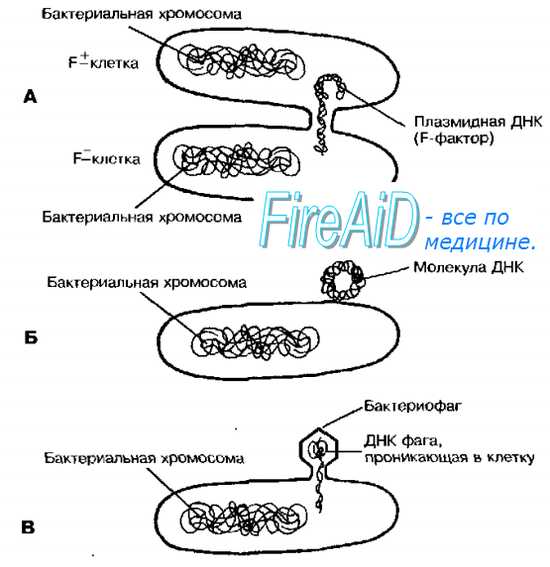 Рис. 4-16. Механизмы перенос бактериальной ДНК. Конъюгация (А), трансформация с использованием отдельной молекулы ДНК (Б) трансдукция с помощью фагов (В).
Рис. 4-16. Механизмы перенос бактериальной ДНК. Конъюгация (А), трансформация с использованием отдельной молекулы ДНК (Б) трансдукция с помощью фагов (В).
Конъюгация бактерий. F-фактор бактерии.
Конъюгация — прямой перенос фрагмента ДНК от донорских бактериальных клеток к реципиентным при непосредственном контакте этих клеток. Биологическая значимость этого процесса стала проясняться после внедрения в медицинскую практику антибиотиков. Устойчивость к антибиотикам можно получить в результате мутации, что происходит один раз на каждые 106 клеточных делений. Однако, однажды изменившись, генетическая информация может быстро распространяться среди сходных бактерий благодаря конъюгации, поскольку каждая третья из близкородственных бактерий способна именно к этому типу генетического переноса. Для реализации процесса необходим F-фактор — плазмида, кодирующая информацию, необходимую для конъюгации.
Конъюгация требует наличия двух типов клеток: доноров (F + ), обладающих F-фактором, и реципиентов (F — ), не обладающих им. При скрещивании клеток F — и F + фактор фертильности передаётся с частотой, близкой к 100%.
Фактор переноса содержит гены специальных и необходимых при конъюгации структур — F-пилей и ряд других генов, вовлечённых в процесс взаимодействия с F — -клетками.
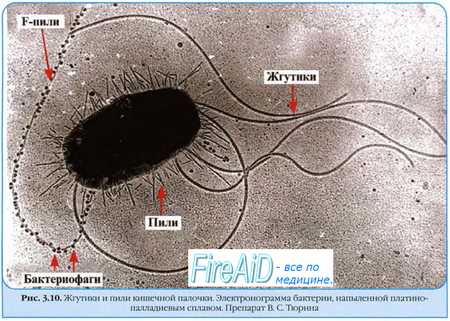
Первый этап конъюгации — прикрепление клетки-донора к реципиенту с помощью F-пилей. Затем между клетками формируется конъюгационный мостик, через который передаётся F-фактор, а также и другие плазмиды, автономно пребывающие в цитоплазме донора. При попадании F-фактора в реципиентную клетку она становится F+ и приобретает способность передавать фактор фертильности другим F — -клеткам. Подобный механизм обеспечивает приобретение популяционно. устойчивости к антибактериальным агентам.
В популяции клеток, содержащих F-плазмиду, только те, в которых она интегрирована в бактериальную хромосому (Hft + -клетки), способны быть донорами хромосомной ДНК. При перенос генетического материала бактериальная ДНК реплицируется, начиная от места включения F-фактора, одна цепь ДНК переносится в реципиентную F’-клетку двигаясь 5′-концом вперёд тогда как другая остаётся в Hfr + -клетке, то есть донор сохраняет своё генетическое постоянство. После начала конъюгации хромосомный материал переносится, начиная от генов, близких к начальной точке транспорта (рис. 4-17).
В бактерии-реципиенты обычно попадают первые из переносимых генов, размер которых зависит от времени, в течение которого проходила конъюгация, и очень редко — все гены. Позже всех переносится участок плазмиды, содержащий ген переноса кодирующий F-пили. Поскольку полная трансмиссия — явление редкое, реципиентная клетка при Hfr-конъюгации обычно остаётся F-. Вслед за процессом переноса в клетке-реципиенте происходит гомологичная рекомбинация между донорской ДНК и собственной ДНК реципиента.
Процесс конъюгации может происходить только при соблюдении ряда условий.
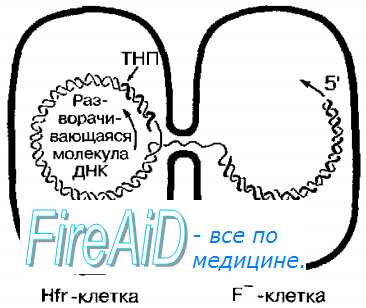 Рис. 4-17. Hfr-конъюгация. Hfr-бактерии функционируют при конъюгации как доноры. Перенос ДНК осуществляется линейно и начинается с удвоения места включения F-фактора (то есть с удвоения точки начала переноса — ТНП).
Рис. 4-17. Hfr-конъюгация. Hfr-бактерии функционируют при конъюгации как доноры. Перенос ДНК осуществляется линейно и начинается с удвоения места включения F-фактора (то есть с удвоения точки начала переноса — ТНП).
• На поверхности реципиентных бактерий должны быть рецепторы пилей, имеющие существенное сродство ( к F-пилям, что позволяет образовать стабильную связь между пилями и рецепторами.
• Для эффективной конъюгации у F-фактора должна быть точка начала репликации, распознаваемая репликативными системами хозяина.
• Эффективность Hfr-конъюгации зависит от величины гомологии ДНК. Перенос негомологичного хромосомного материала донора не приведёт к его интеграции с ДНК реципиента.
Источник
Бактериальная конъюгация — Bacterial conjugation
Бактериальная конъюгация — это передача генетического материала между бактериальными клетками посредством прямого межклеточного контакта или мостикового соединения между двумя клетками. Это происходит через пилус . Это парасексуальный способ размножения бактерий.
Это механизм горизонтального переноса генов, а также трансформация и трансдукция, хотя эти два других механизма не связаны с межклеточным контактом.
Классическая бактериальная конъюгация E. coli часто рассматривается как бактериальный эквивалент полового размножения или спаривания, поскольку она включает обмен генетическим материалом. Однако это не половое размножение, так как не происходит обмена гамет и, по сути, не происходит генерации нового организма : вместо этого трансформируется существующий организм. Во время классической конъюгации E. coli донорская клетка обеспечивает конъюгированный или мобилизуемый генетический элемент, который чаще всего является плазмидой или транспозоном . Большинство конъюгативных плазмид имеют системы, гарантирующие, что клетка- реципиент еще не содержит подобный элемент.
Передаваемая генетическая информация часто приносит пользу получателю. Преимущества могут включать устойчивость к антибиотикам , толерантность к ксенобиотикам или способность использовать новые метаболиты . Другие элементы могут быть вредными и могут рассматриваться как бактериальные паразиты .
Конъюгация у Escherichia coli посредством спонтанного зигогенеза и у Mycobacterium smegmatis посредством дистрибутивного конъюгированного переноса отличается от лучше изученной классической конъюгации E. coli тем, что в этих случаях происходит существенное смешение родительских геномов .
СОДЕРЖАНИЕ
История
Этот процесс был открыт Джошуа Ледербергом и Эдвардом Татумом в 1946 году.
Механизм
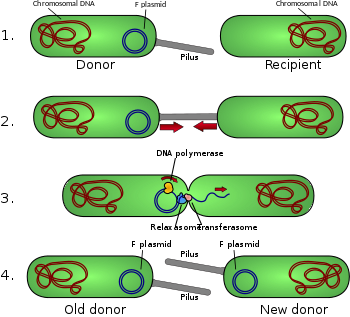
- Донорская клетка производит пилус .
- Пилус прикрепляется к реципиентной клетке и объединяет две клетки.
- Подвижная плазмида разрывается, и затем одиночная цепь ДНК переносится в реципиентную клетку.
- Обе клетки синтезируют комплементарную цепь для получения двухцепочечной кольцевой плазмиды, а также воспроизводят пили; обе клетки теперь являются жизнеспособными донорами F-фактора.
F-плазмида является эписомой (плазмида , которая может интегрироваться в бактериальную хромосому пути гомологичной рекомбинации ) с длиной около 100 кб . Он несет свою собственную точку начала репликации , oriV , и точку начала передачи, или oriT . В данной бактерии может быть только одна копия F-плазмиды, свободная или интегрированная, а бактерии, обладающие такой копией, называются F-позитивными или F-плюс (обозначаются F + ). Клетки, в которых отсутствуют плазмиды F, называются F-отрицательными или F-отрицательными (F — ) и как таковые могут функционировать как клетки-реципиенты.
Помимо другой генетической информации, F-плазмида несет локусы tra и trb , которые вместе имеют длину около 33 т.п.н. и состоят из около 40 генов . TRA локус включает пилина ген и регуляторные гены, которые вместе образуют фимбрии на клеточной поверхности. Локус включает также гены для белков , которые прикрепляются к поверхности F — бактерии и инициировать сопряжение. Хотя есть некоторые споры о точном механизме конъюгации, кажется, что пили не являются структурами, через которые происходит обмен ДНК. Это было показано в экспериментах, в которых пилусам позволяют контактировать, но затем они денатурируются с помощью SDS, но трансформация ДНК все еще продолжается. Некоторые белки, кодируемые в локусе tra или trb, по- видимому, открывают канал между бактериями, и считается, что фермент traD, расположенный в основании пилуса, инициирует слияние мембран.
Когда конъюгация инициируется сигналом, фермент релаксаза создает разрыв в одной из цепей конъюгативной плазмиды в oriT . Релаксаза может работать самостоятельно или в комплексе из более чем дюжины белков, известных под общим названием релаксосома . В системе F-плазмиды фермент релаксазы называется TraI, а релаксосома состоит из TraI, TraY, TraM и интегрированного фактора хозяина IHF. Разорванная цепь или Т-цепь затем разматывается с неразрывной цепи и переносится в реципиентную клетку в направлении от 5′-конца к 3′-концу. Остальные нити реплицируются либо не зависят от конъюгативного действия (вегетативной репликации , начиная с oriV ) или совместно с сопряжением (конъюгативной репликацией аналогично прокатной окружностью репликации фага лямбды ). Для конъюгативной репликации может потребоваться второй ник, прежде чем может произойти успешный перенос. В недавнем отчете утверждается, что он ингибировал конъюгацию с химическими веществами, которые имитируют промежуточный этап этого второго события пощипывания.
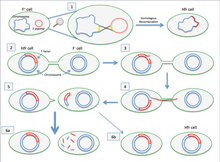
Если переносимая F-плазмида была ранее интегрирована в геном донора (продуцируя штамм Hfr [«Высокая частота рекомбинации»]), часть хромосомной ДНК донора также может быть перенесена с плазмидной ДНК. Количество передаваемой хромосомной ДНК зависит от того, как долго две конъюгированные бактерии остаются в контакте. В обычных лабораторных штаммах E. coli перенос всей бактериальной хромосомы занимает около 100 минут. Затем перенесенная ДНК может быть интегрирована в геном реципиента посредством гомологичной рекомбинации .
Культура клеток, которая содержит в своей популяции клетки с не интегрированными F-плазмидами, обычно также содержит несколько клеток, которые случайно интегрировали свои плазмиды. Именно эти клетки ответственны за низкочастотный перенос хромосомных генов, происходящий в таких культурах. Некоторые штаммы бактерий с интегрированной F-плазмидой можно выделить и выращивать в чистой культуре. Поскольку такие штаммы передача хромосомных генов очень эффективно они называется Hfr ( ч IGH е requency из г ecombination). E.coli , геном был первоначально нанесен на карту прерванных экспериментов спаривания , в которых различные клетки Hfr в процессе конъюгации были срезанными от получателей после менее чем за 100 минут (первоначально с использованием блендер Waring). Затем были исследованы переданные гены.
Поскольку интеграция F-плазмиды в хромосому E. coli является редким спонтанным явлением, и поскольку многочисленные гены, способствующие переносу ДНК, находятся в геноме плазмиды, а не в геноме бактерий, утверждалось, что конъюгативный перенос бактериальных генов, как он встречается в системе Hfr E. coli , не является эволюционной адаптацией бактериального хозяина и, вероятно, не является предком эукариотического пола.
Спонтанный зигогенез у E. coli
В дополнение к классической бактериальной конъюгации, описанной выше для E. coli , в некоторых штаммах E. coli наблюдается форма конъюгации, называемая спонтанным зигогенезом (Z-спаривание для краткости) . При Z-спаривании происходит полное генетическое смешение, и образуются нестабильные диплоиды , которые отбрасывают фенотипически гаплоидные клетки, некоторые из которых демонстрируют родительский фенотип, а некоторые являются истинными рекомбинантами .
Конъюгальный перенос микобактерий
Конъюгация у Mycobacteria smegmatis , как и конъюгация в E. coli , требует стабильного и продолжительного контакта между донором и штаммом-реципиентом, устойчива к ДНКазе, а перенесенная ДНК включается в хромосому реципиента путем гомологичной рекомбинации. Однако, в отличие от конъюгации Hfr E. coli , микобактериальная конъюгация основана на хромосоме, а не плазмиде. Кроме того, в отличие от конъюгации Hfr E. coli , у M. smegmatis все области хромосомы переносятся с сопоставимой эффективностью. Длина донорских сегментов сильно различается, но средняя длина составляет 44,2 КБ. Поскольку в среднем переносится 13 участков, среднее количество перенесенной ДНК на геном составляет 575 КБ. Этот процесс называется «распределительный супружеский перенос». Gray et al. обнаружили существенное смешение родительских геномов в результате конъюгации и считали, что это смешение напоминает то, что наблюдается в мейотических продуктах полового размножения.
Передача между королевствами

Бактерии , связанные с фиксацией азота ризобий представляют собой интересный случай интер- царства сопряжения. Например, индуцирующая опухоль (Ti) плазмида Agrobacterium и плазмида, индуцирующая корневую опухоль (Ri) A. rhizogenes, содержат гены, которые способны переноситься в клетки растений. Экспрессия этих генов эффективно преобразовывает растительные клетки в опин -продуцирующих заводов. Опины используются бактериями как источники азота и энергии. Зараженные клетки образуют коронковый галл или корневую опухоль . Таким образом, плазмиды Ti и Ri являются эндосимбионтами бактерий, которые, в свою очередь, являются эндосимбионтами (или паразитами) инфицированного растения.
Плазмиды Ti и Ri могут также передаваться между бактериями с использованием системы ( tra , или перенос, оперон ), которая отличается и не зависит от системы, используемой для передачи между царствами ( vir , или вирулентность , оперон). Такие передачи создают вирулентные штаммы из ранее невирулентных штаммов.
Приложения генной инженерии
Конъюгация — удобный способ передачи генетического материала различным мишеням. В лабораториях сообщалось об успешном переносе бактерий в дрожжи, растения, клетки млекопитающих, диатомовые водоросли и изолированные митохондрии млекопитающих . Конъюгация имеет преимущества по сравнению с другими формами генетического переноса, включая минимальное нарушение клеточной оболочки- мишени и способность переносить относительно большие количества генетического материала (см. Выше обсуждение переноса хромосомы E. coli ). В растениеводстве конъюгация, подобная Agrobacterium, дополняет другие стандартные носители, такие как вирус табачной мозаики (TMV). Хотя TMV способен инфицировать многие семейства растений, это в первую очередь двудольные травянистые растения . Конъюгация, подобная Agrobacterium , также в основном используется для двудольных, но реципиенты однодольных не редкость.
Источник



