
рабочая тетрадь клетка. Материалы для подготовки к занятию Лекция. Введение в физиологию клетки Материалы для подготовки Строение клетки
| Название | Материалы для подготовки к занятию Лекция. Введение в физиологию клетки Материалы для подготовки Строение клетки |
| Дата | 18.03.2019 |
| Размер | 1.1 Mb. |
| Формат файла |  |
| Имя файла | рабочая тетрадь клетка.docx |
| Тип | Материалы для подготовки #70798 |
| страница | 5 из 5 |
| Подборка по базе: Задание для сам. подготовки по теме №3 (2).docx, Темы по биологии, для подготовки к медицинскому турниру Предунив, русский язык и культура речи учебные материалы02.doc, контрольно-и змерительные материалы_copy.pdf, Тестовые задания для подготовки к итоговой государственной аттес, Вопросы для подготовки к итоговому тестированию по дисциплине.do, Практические задания к семинарскому занятию10.docx, Инструкция по подключению к онлайн занятию.docx, ЧЕК-ЛИСТ подготовки к ЕГЭ.pdf, Контрольные вопросы для самостоятельной подготовки.docx Работа № Роль нейромышечного синапса в возникновении утомления Виртуальный практикум «LupraFisim» В возникновении сокращения скелетных мышц задействованы три структуры: 1 двигательный нейрон; 2 нейромышечный синапс; 3 волокно скелетной мышцы. Из этих трех структур только моторный нейрон не подвержен явлению утомления, его практически не возможно утомить. В двух других структурах возможно возникновение утомления. Цель: продемонстрировать, что нейромышечный синапс утомляется быстрее, чем мышечное волокно. Принцип действия: на двигательный нейрон поперечно-полосатой мышцы воздействуют залпом электрических стимулов, одновременно получая миограмму, до тех пор, пока мышца не перестанет сокращаться (проявляется утомление). Затем раздражитель перемещают на саму мышцу и подвергают воздействию стимулов уже непосредственно ее. — с помощью соответствующих кнопок выберите вариант «НЕПРЯМОЙ СТИМУЛ» (воздействию стимула подвергается двигательный нерв, а непосредственно мышца); — щелкнув по кнопке «ВОЗДЕЙСТВОВАТЬ ПАЧКОЙ СТИМУЛОВ», начните воздействовать на мышцу группой стимулов; — внимательно наблюдайте за изменениями, которые претерпевает миограмма; заметьте, что с течением времени амплитуда сокращений постепенно уменьшается; — после того, как можно констатировать, что мышца более не сокращается (проявляется утомление) при продолжающемся воздействии на нее залпов стимулов, измените способ воздействия, для чего, щелкнув по соответствующей кнопке, выберите вариант «ПРЯМОЙ СТИМУЛ» (то есть стимул, воздействующий непосредственно на мышцу); — анализируя полученную миограмму, следует констатировать, что, с началом воздействия раздражителем непосредственно на мышцу, она начинает сокращаться снова (призрак того, что утомления в самой мышце еще не возникло, а утомление, проявившееся прежде, возникло из-за утомления нейромышечного синапса), впрочем, с амплитудой несколько меньшей, которая постепенно уменьшается и уменьшается и далее, пока мышца не перестанет сокращаться (возникает собственное мышечное утомление). При непрямом воздействии на мышечное волокно происходят сокращения мышцы, причем с течением времени их амплитуда уменьшается вплоть до полного прекращения. При переключении на прямое стимулирование мышца вновь начинает сокращаться примерно так же, как и в первом случае. Раньше всего утомление наступает в зоне передачи сигнала от нерва к мышце – синаптической бляшке. Далее при переключении на непосредственное стимулирование мышцы происходит и ее утомление. Нервная ткань более склонна к утомлению. Занятие №7 Материалы для подготовки к занятию: Лекция. Молекулярные механизмы нейрогуморальной регуляции 1. Дать определение понятия регуляции функции. Регуляция – управление физиологическими функциями, деятельностью клеток, тканей, органов, систем, поведением организма, осуществление взаимодействия организма и окружающей среды 3. Дать определение гуморального механизма регуляции. Гуморальный механизм регуляции -это один из эволюционно ранних механизмов регуляции процессов жизнедеятельности в организме, осуществляемый через жидкие среды организма (кровь, лимфу, тканевую жидкость, слюну) с помощью гормонов, выделяемых клетками, органами, тканями 4. Дать сравнительную характеристику нервного и гуморального механизмов регуляции
5. Перечислите основные группы факторов гуморальной регуляции Факторы гуморальной регуляции (группы веществ, участвующие в гуморальной регуляции) 1. Неорганические метаболиты и ионы. Например, катионы кальция, водорода, углекислый газ. 2. Гормоны желез внутренней секреции. Вырабатываются специализированными инкреторными железами. Это инсулин, тироксин и др. 3. .Местные или тканевые гормоны. Эти гормоны вырабатываются специальными клетками, называемыми паракринными, транспортируются тканевой жидкостью и действуют только на небольшом расстоянии от секретирующих клеток. К ним относятся такие вещества, как гистамин, серотонин, гормоны желудочно-кишечного тракта и другие. 4. Биологически активные вещества, обеспечивающие креаторные связи между клетками ткани. Это белковые макромолекулы, выделяемые ими. Они регулируют дифференцировку, рост и развитие всех клеток составляющих ткань и обеспечивают функциональное объединение клеток в ткань. Такими белками являются, например, кейлоны, которые тормозят синтез ДНК и деление клеток. 6. Перечислите способы доставки сигнальных молекул к клетке-мишени при гуморальном механизме регуляции. 2. Взаимодействие молекул клеточных мембран 3. Передача сигнальных молекул через межклеточное пространство 7. Понятие об аутокринном способе передачи сигнальных молекул. аутокринный способ — это способ передачи сигнала при котором продуцент гормона имеет рецепторы к этому же гормону (другими словами, клетка—продуцент гормона в то же время является его мишенью). Примеры: эндотелины, вырабатываемые клетками эндотелия и воздействующие на эти же эндотелиальные клетки; Т-лимфоциты, секретирующие интерлейкины, имеющие мишенями разные клетки, в том числе и Т-лимфоциты 8. Понятие о паракринном способе передачи сигнальных молекул. паракринный способ — это способ передачи сигнала при котором продуцент биологически активного вещества и клетка-мишень расположены рядом. Молекулы гормона достигают мишени путём диффузии в межклеточном веществе. Например, в париетальных клетках желёз желудка секрецию Н+ стимулируют гастрин и гистамин, а подавляют соматостатин и Пг, секретируемые рядом расположенными клетками. 9. Понятие о телекринном способе передачи сигнальных молекул. телекринный способ – это способ передачи сигнальных молекул непосредственно от наружной поверхности мембраны одной клетки на мембрану другой. Это происходит при условии непосредственного контакта (прикрепления, адгезионного сцепления) мембран двух клеток. Такое прикрепление происходит, например, при взаимодействии лейкоцитов и тромбоцитов с эндотелием кровеносных капилляров в месте, где имеется воспалительный процесс 10. Перечислите принципы регуляции функции. 1. Для нормальной регуляции функций организма необходимо взаимодействие нервной и гуморальной систем. 2. Нейрогуморальная регуляция объединяет все функции организма для достижения цели, при этом организм функционирует как единое целое. Обратная связь это процесс, приводящий к тому, что результат функционирования какой-либо системы влияет на параметры, от которых зависит функционирование этой системы. 12. Виды обратной связи:
Положительная обратная связь – это тип обратной связи, при котором изменение выходного сигнала системы приводит к такому изменению входного сигнала, которое способствует дальнейшему отклонению выходного сигнала от первоначального значения.
Отрицательная обратная связь – это вид обратной связи, при котором изменение выходного сигнала системы приводит к такому изменению входного сигнала, которое противодействует первоначальному изменению Вопросы для самоконтроля: 1. Понятие о регуляции функции, ее значение для адаптации. 2. Понятие о механизме регуляции функции. Основные механизмы регуляции с краткой сравнительной характеристикой. 3. Понятие о гуморальном механизме регуляции, основные факторы гуморальной регуляции. 4. Понятие о гормональной регуляции. Понятие гормона, их биологическое значение. 5. Классификация и основные свойства гормонов. 6. Секреция, транспорт, инактивация гормонов. 7. Основные принципы регуляции функции. 8. Понятие обратной связи. Положительная и отрицательная обратная связь. Работа №1. Воздействие медикаментов и медиаторов на деятельность сердца. Виртуальный практикум «LupraFisim» Пронаблюдать возможность влияния гуморальных факторов на изолированное сердце лягушки. Оборудование: Р-ры KCl, CaCl2, адреналина. Объект исследования: Лягушка. Ход работы: Подсчитать частоту сокращений изолированного сердца лягушки до и после добавления растворов электролитов и адреналина. Результаты занести в таблицу:
Действие химических медиаторов на изолированное сердце несущественно влияет на изменение частоты ЧСС. Тема: Физиология клетки. Механизмы межклеточного взаимодействия Цель:
Алгоритм проведения зачетного занятия. 1. Тестирование, каждый студент получает задание из 50 тестов по всем модулям дисциплины. «Зачтено» если студент суммарно набрал не менее 70% правильных ответов. Источник Аутокринный способ это физиологияа) Высвобождение медиаторов. В состоянии покоя синаптические пузырьки накапливаются в активных зонах нервных окончаний и направляются к пресинаптическим уплотнениям при участии докинг-белков, один из которых — актин. При прохождении потенциала действия происходит открытие потенциалозависимых кальциевых (Са 2+ ) каналов, расположенных в непосредственной близости к активной зоне пресинаптической мембраны, что приводит к быстрому заполнению этой области ионами Са 2+ . Ионы Са 2+ связываются с определенными белками, расположенными в области скопления синаптических пузырьков и в активной зоне, в результате чего происходят слияние синаптических пузырьков с клеточной мембраной и высвобождение нейромедиаторов. Процесс слияния синаптических пузырьков с клеточной мембраной начинается с образования плотного комплекса между везикулярными белками SNARE (v-SNARE) и белками пресинаптической мембраны (t-SNARES). Метаболические компоненты, необходимые для осуществления слияния синаптических пузырьков с мембраной клетки, расположены в активной зоне. Наиболее важный крупный мультидоменный белок RIM соединяется с ГТФ-связывающим белком синаптических пузырьков Rab3. Остальные белки обеспечивают слияние синаптических пузырьков с мембраной клетки и высвобождение нейромедиаторов. К синаптическим компонентам относят синаптические везикулярные белки (синаптотагмин, синаптобревин, синаптофизин и синапсины), везикуло-ассоциированные белки (амфифизин, динамин и СаМ-киназы), синаптические белки плазматической мембраны (синтаксины, нейрексины и SNAP-25), а также цитозольные белки (комплексины, различные виды белков SNAP и NSF-белки). Многие из перечисленных синаптических белков выполняют специфические функции по обеспечению единства процессов возбуждения клетки и высвобождения нейромедиаторов, а также механизмов восстановления синаптических пузырьков, что лежит в основе синаптической передачи сигнала. Молекулярные процессы, обеспечивающие взаимосвязь возбуждения клетки и высвобождения нейромедиаторов, представляют собой сложные модели, осуществляемые различными механизмами. Белки сразу же формируют ответную реакцию на поступление ионов Са 2+ в клетку, в связи с чем временной промежуток между входом ионов и высвобождением нейромедиаторов составляет менее 1 мс. Для высвобождения некоторого количества нейромедиаторов из синаптических пузырьков небольшого размера [например, содержащих глутамат или у-аминомасляную кислоту (ГАМК)] достаточно единичных потенциалов действия. В это же время для запуска медленного (с задержкой 50 мс и выше) высвобождения медиаторов из гранулярных везикул большого размера, характерных для пептидергических нейронов, минимальная частота импульсов составляет 10 Гц. Таким образом, количество высвобождаемого медиатора не является постоянной величиной и может изменяться под влиянием внутренних и внешних факторов. б) Связывание медиатора с рецептором клетки-мишени. Молекулы нейромедиатора связываются с молекулами белков-рецепторов на постсинаптической мембране. Выделяют ионотропные и метаботропные рецепторы. Каждая группа включает рецепторы, активация которых приводит к открытию ионных каналов, а также рецепторы, обеспечивающие закрытие этих каналов. 1. Ионотропные рецепторы. В каждой макромолекуле ионотропных рецепторов присутствует ионный канал. Медиатор связывается со специфическим рецептором в синаптической щели, вызывая ее конформационное преобразование, что приводит к открытию ионного канала. Считают, что ионотропные каналы являются медиаторозависимыми (или лиганд-зависимыми), что указывает на их способность связываться с молекулой медиатора или лекарственного вещества. После отсоединения молекулы нейромедиатора или его разрушения ионный канал, закрываясь, возвращается в исходное состояние. Нейромедиатор вызывает открытие возбуждающего ионного канала, в результате чего происходят значительное поступление ионов натрия (Na + ) в клетку и незначительный выход ионов калия (К + ) из клетки. Эти процессы приводят к образованию возбуждающего постсинаптического потенциала (ВПСП), вызывающего деполяризацию мембраны. При открытии множества медиаторозависимых каналов происходит суммирование возбуждения и возникает деполяризация большей силы, которая может достигать пороговых значений и вызывать образование потенциала действия. Тормозной постсинаптический потенциал (ТПСП), следующий за ВПСП, вызывает гиперполяризацию мембраны до -70 мВ, что служит равновесным потенциалом для ионов хлора (Cl — ). При открытии калиевых каналов, равновесный потенциал ионов которых составляет -80 мВ, происходит гиперполяризация большей силы. Ионотропные рецепторы получили название «быстрых», поскольку они оказывают быстрое, но непродолжительное действие на ионные каналы. 2. Метаботропные рецепторы. Метаботропные рецепторы получили свое название благодаря способности оказывать в цитоплазме нейрона многочисленные метаболические действия. Макромолекула рецептора представляет собой трансмембранный белок, в котором отсутствует ионный канал. Рецептор активируется посредством присоединения медиатора к его активному центру, в результате чего происходит конформационная перестройка белка, что, в свою очередь, активирует одну из закрепленных субъединиц (α- или β-субъединицу). После активации субъединица открепляется и направляется вдоль внутренней поверхности клеточной мембраны. Субъединицы являются G-белками за счет преимущественного связывания с гуанинтрифосфатом (ГТФ) или гуаниндифосфатом (ГДФ). В большинстве случаев эти белки оказывают непрямое действие через систему вторичных посредников. Однако некоторые G-белки активируют ионные каналы напрямую. G-белок, оказывающий возбуждающее действие, называют Gs-белком, а белок, оказывающий тормозное действие,— Gi-белком. За счет многостадийного функционирования метаботропные рецепторы, как правило, являются «медленными»; после единичного стимула влияние мембранного канала может продолжаться сотни миллисекунд. Кроме того, образование вторичных посредников может привести к снижению способности нейрона к возбуждению. В настоящее время выделяют три системы вторичных посредников. 1. Система циклического аденозинмонофосфата (цАМФ), осуществляющая фосфорилирование белков. 2. Инозитолфосфатная система, обеспечивающая высвобождение ионов Са2+ из эндоплазматических депо. 3. Система арахидоновой кислоты, за счет которой осуществляется образование метаболитов арахидоновой кислоты. — Система цАМФ. Связывание нейромедиатора с рецептором приводит к отделению а-субъединицы от Gs-белка, за счет чего становится возможным присоединение ГТФ к освободившемуся белку, который, в свою очередь, способствует превращению аденозинтрифосфата (АТФ) в цАМФ под действием аденилатциклазы (рис. 8.4). Синтезированный цАМФ, расщепляясь, выполняет в клетке функцию вторичного посредника. Под воздействием цАМФ протеинкиназа А переносит фосфат-ионы от молекулы АТФ в ионный канал, что приводит к его открытию и поступлению ионов Na + внутрь клетки, за счет чего происходит деполяризация нейрона-мишени. При инактивации Gs-белка под действием прикрепленного к мембране фермента протеинфосфатазы происходят выход фосфат-ионов из клетки и закрытие ионного канала. — Инозитолфосфатная система. Эффекторный фермент — фосфолипаза С — под влиянием другой активированной а-субъединицы G -белка расщепляет фосфолипид мембраны PIP, с образованием двух вторичных посредников; диацилглицерола (ДАГ) и инозитолтрифосфата (ИТФ). ДАГ активирует протеинкиназу С, которая запускает процесс фосфорилирования белков. ИТФ, проникая в цитозоль, открывает кальций-зависимые ионные каналы преимущественно вблизи мембран гладкой эндоплазматической сети. Ионы Са 2+ запускают Са 2+ -зависимый каскад ферментов, осуществляющих открытие и/или закрытие ионных каналов, а также, вероятно, изменяющих экспрессию генов и синтез белков при прохождении через ядерную оболочку (см. далее — транскрипция генов). Система арахидоновой кислоты. Описана далее (в связи с гистамином). — Транскрипция генов. В настоящее время установлено, что рефлекторный ответ на повторяющиеся воздействия способен либо прогрессивно увеличиваться при формировании сенсибилизации, что в большинстве случае происходит под влиянием вредоносных стимулов, либо уменьшаться при формировании приспособления, что чаще встречается при воздействии безвредных стимулов. В ходе экспериментов на животных, изучались рефлекторные дуги, в которых были задействованы чувствительные, двигательные и ассоциативные нейроны, было показано, что для развития сенсибилизации характерно образование новых синаптических контактов между ассоциативными и двигательными нейронами; это сопровождается дополнительным синтезом и высвобождением медиаторов. Для развития приспособления, напротив, характерно снижение синтеза и высвобождение медиаторов. Данные процессы обусловлены изменением транскрипции генов. Повторяющиеся вредоносные стимулы оказывают влияние на цАМФ, в результате чего происходит чрезмерная активация протеинкиназ, участвующих в фосфорилировании белков, регулирующих транскрипцию генов. При этом увеличивается синтез белков (в том числе и ферментов), необходимых для синтеза медиаторов и построения новых ионных каналов и цитоскелета синапса. Действие повторяющихся безвредных стимулов заключается только в снижении синтеза и высвобождения медиаторов. Влияния транскрипции генов играют особенно значимую роль в процессе формирования долгосрочной памяти. Видео физиология синапса и нерва — профессор, д.м.н. П.Е. УмрюхинРедактор: Искандер Милевски. Дата публикации: 11.11.2018 Источник |
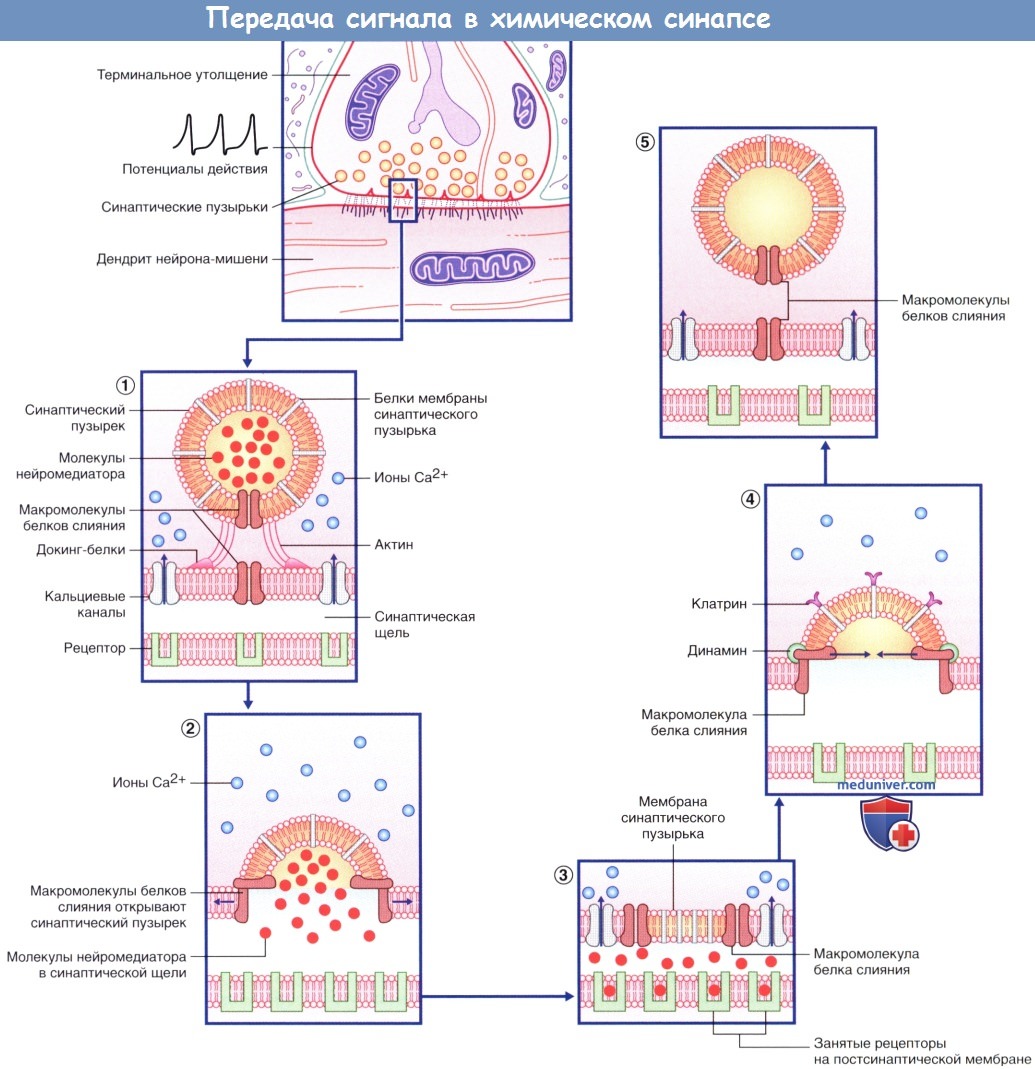 Последовательность процессов, происходящих в клетке после деполяризации пресинаптической мембраны.
Последовательность процессов, происходящих в клетке после деполяризации пресинаптической мембраны. 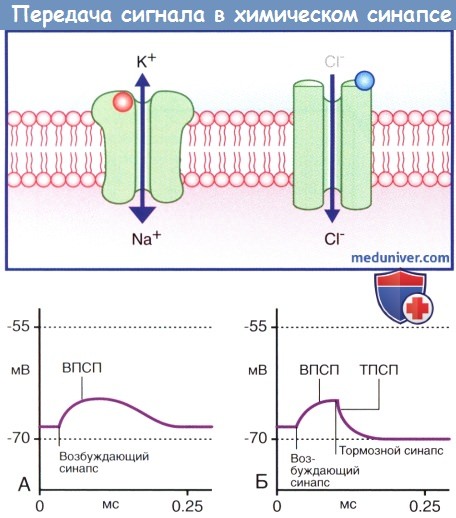 (А) Медиаторозависимый возбуждающий ионотропный рецептор.
(А) Медиаторозависимый возбуждающий ионотропный рецептор. 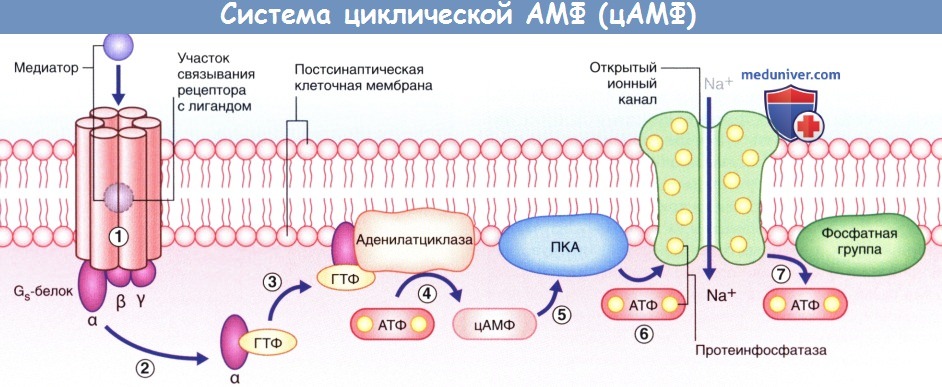 Система циклической АМФ (цАМФ).
Система циклической АМФ (цАМФ). 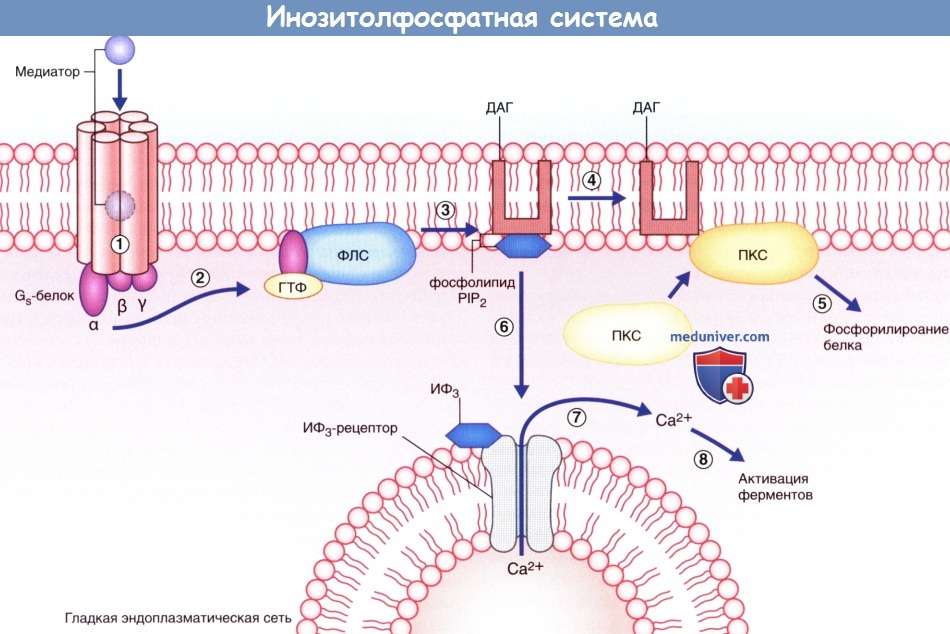 Инозитолфосфатная система.
Инозитолфосфатная система. 


