
- Анаэробное дыхание
- Из Википедии — свободной энциклопедии
- Анаэробное дыхание
- Типы анаэробного дыхания
- Метаболические особенности анаэробных бактерий
- Анаэробный способ дыхания у кого
- Анаэробное дыхание
- Содержание
- Нитратное и нитритное дыхание
- Сульфатное дыхание
- Фумаратное дыхание
- Железистое дыхание
- Дыхание ацетогенных бактерий
- Анаэробное дыхание у растений
- Анаэробное дыхание у грибов, протистов и животных
Анаэробное дыхание
Из Википедии — свободной энциклопедии
Анаэробное дыхание — это биохимический процесс окисления органических субстратов или молекулярного водорода с использованием в дыхательной ЭТЦ в качестве конечного акцептора электронов вместо O2 других окислителей неорганической или органической природы. Как и в случае аэробного дыхания, выделяющаяся в ходе реакции свободная энергия запасается в виде трансмембранного протонного потенциала, использующегося АТФ-синтазой для синтеза АТФ.
Осуществляется прокариотами (в редких случаях — и эукариотами) в анаэробных условиях. При этом факультативные анаэробы используют акцепторы электронов с высоким окислительно-восстановительным потенциалом (NO3 − , NO2 − , Fe 3+ , фумарат, диметилсульфоксид и т. д.), у них это дыхание конкурирует с энергетически более выгодным аэробным и подавляется кислородом. Акцепторы с низким окислительно-восстановительным потенциалом (сера, SO4 2− , CO2) применяются только строгими анаэробами, гибнущими при появлении в среде кислорода. В корневых системах многих растений при гипоксии и аноксии, вызванных затоплением посевов в результате длительных дождей или весенних паводков, развивается анаэробное дыхание с использованием в качестве акцепторов электронов альтернативных кислороду соединений, например нитратов. Установлено, что растения, произрастающие на полях, удобренных нитратными соединениями, переносят переувлажнение почвы и сопутствующую ему гипоксию лучше, нежели такие же растения без нитратной подкормки.
Механизмы окисления органических субстратов при анаэробном дыхании, как правило, аналогичны механизмам окисления при аэробном дыхании. Исключением является использование в качестве исходного субстрата ароматических соединений. Обычные пути их катаболизма требуют молекулярного кислорода уже на первых стадиях, в анаэробных условиях осуществляются иные процессы, например, восстановительная деароматизация бензоил-КоА у Thauera aromatica с затратой энергии АТФ. Некоторые субстраты (например, лигнин) при анаэробном дыхании не могут использоваться.
Источник
Анаэробное дыхание

Анаэробное дыхание – это такое дыхание микробов, при котором для окисления органических или неорганических веществ используется не молекулярный кислород, а другие окисленные соединения. В частности, соли азотной, серной и угольной кислот. При этом используемые соединения превращаются в более восстановленные. Процессы идут в анаэробных условиях, то есть без доступа кислорода [1] .
При анаэробном дыхании акцептором электронов выступают различные неорганические соединения – нитраты, сульфаты, карбонаты. Таким образом, различия между аэробным дыханием и анаэробным дыханием заключено в природе конечного акцептора электронов. Выход аденозинтрифосфорной кислоты (АТФ) при анаэробном дыхании меньше, чем при аэробном, но больше, чем при брожении [2] [1] .
Типы анаэробного дыхания
Тип анаэробного дыхания зависит от природы конечного акцептора электронов в электротранспортной цепи. В зависимости от этого различают:
- сульфатное дыхание – конечный акцептор электронов сульфатная группа SO4 2- . Бактерии – сульфатвосстанавливающие или сульфатредуцирующие;
- нитратное дыхание (денитрификация) – конечный акцептор электронов нитратная (NO3 — ) или нитритная группа (NO2 — ). Бактерии – денитрифицирующие;
- карбонатное дыхание – конечный акцептор электронов CO2 (улекислый газ). Бактерии – метаногенные (метанобразующие);
- фумаратное дыхание – конечным акцептором электронов служит органическое вещество [2] .
Метаболические особенности анаэробных бактерий
Бактерии способные к анаэробному дыханию характеризуются укороченными электротранспортными или дыхательными цепями. Они не содержат всех переносчиков, характерных для дыхательных цепей аэробных бактерий [2] .
Параллельно в цепях анаэробов цитохромоксидаза заменена соответствующими редуктазами. В обмене веществ строгих анаэробов не функционирует цикл Кребса или он разорван и выполняет только биосинтетические, но не энергетические функции [2] .
Основное количество молекул АТФ при анаэробном дыхании синтезируется в процессе мембранного фосфорилирования [2] .
По отношению к молекулярному кислороду бактерии, для которых характерно анаэробное дыхание, могут являться факультативными или облигатными анаэробами [2] .
Облигатные анаэробы – это сульфатвосстанавливающие и метаногенные бактерии [2] .
Факультативные анаэробы – это денитрифицирующие бактерии и бактерии, осуществляющие фумаратное дыхание [2] .
Факультативные анаэробы могут переключать свой энергетический метаболизм с аэробного дыхания в присутствии в среде молекулярного кислорода, на анаэробное дыхание в отсутствии молекулярного кислорода [2] .
Источник
Анаэробный способ дыхания у кого
Анаэробное дыхание часто называют брожением. Многие микроорганизмы получают большую часть своего АТФ за счет анаэробного дыхания. Для некоторых бактерий кислород, даже в обычных присутствующих в атмосфере количествах, вообще губителен, так что они вынуждены жить там, где нет кислорода. Такие организмы называют облигатными анаэробами (пример — Clostridium tetani, возбудитель столбняка).
Известны и другие организмы, например дрожжи и паразиты кишечного тракта (ленточные черви и др.), которые могут существовать как без кислорода, так и и его присутствии. Их называют факультативными анаэробами: при необходимости они переходят на анаэробное дыхание, однако в присутствии кислорода используют аэробный путь. Некоторые клетки, временно испытывающие недостаток кислорода (в частности, мышечные клетки), также обладают способностью к анаэробному дыханию.
Первой фазой анаэробного дыхания тоже является гликолиз. Он дает в результате на каждую молекулу глюкозы две молекулы пировиноградной кислоты, две молекулы АТФ и две молекулы восстановленного НАД (см. табл. 9.1). При аэробном дыхании присоединившийся к НАД водород после ряда реакций, идущих с высвобождением энергии, передается в конце концов кислороду и окисляется до воды. При анаэробном дыхании это оказывается невозможным, поскольку кислорода нет. Вместо этого водород вновь присоединяется к пировиноградной кислоте, так что часть энергии, заключенной в молекуле глюкозы, так и не извлекается (остается в конечном продукте брожения). Ниже мы подробнее рассмотрим, как это происходит у грибов и в животных клетках.
Анаэробное дыхание у грибов, например у дрожжей
Пировиноградная кислота———> Ацетальдегид + СО2
Фермент: пируватдекарбоксилаза
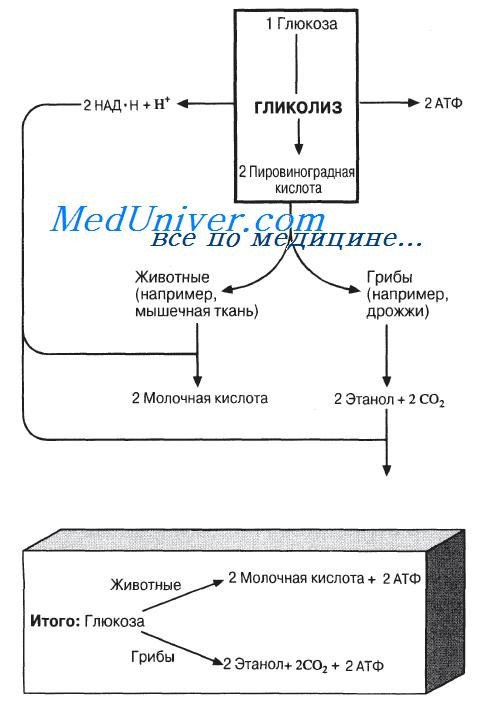
Ацетальдегид + НАД • Н + Н+———> Этанол + НАД+
Фермент: алкогольдегидрогеназа
Итого: Пировиноградная кислота———> Этанол + СO2
Здесь приведены конечные этапы процесса, который носит название спиртового брожения. АТФ при спиртовом брожении образуется только на ранних его этапах — при расщеплении глюкозы до пировиноградной кислоты. Спиртовое брожение используется в производстве пива, вина и других спиртных напитков. В производстве хлебобулочных изделий используют выделяемый дрожжами в процессе спиртового брожения СОг — пузырьки этого газа заставляют подниматься тесто. Конечный продукт спиртового брожения — этанол — содержит еще довольно много энергии (в Бразилии, например, из него делают газохол, на котором ездят автомобили). Однако в отсутствие кислорода энергию из этанола извлечь нельзя.
Общий выход АТФ при спиртовом брожении составляет две молекулы АТФ на одну молекулу глюкозы.
Анаэробное дыхание в животных клетках, например в мышечной ткани
Пировиноградная кислота + НАД * Н + Н+ —————-> Молочная кислота + НАД*
Фермент: лактатдегидрогеназа
В отличие от спиртового брожения ни СО2, ни этанол при молочнокислом брожении не образуются. Конечным продуктом в данном случае является молочная кислота, накопление которой в мышцах вызывает чувство усталости, а иногда и судороги. О кислородной задолженности, возникающей при усиленной мышечной работе, мы будем говорить в следующих статьях.
При молочнокислом брожении, так же как и при спиртовом, на одну молекулу глюкозы образуются две молекулы АТФ. В конечном его продукте — молочной кислоте — сохраняется еще много энергии.
Общая схема анаэробного дыхания приведена на рисунке.
Источник
Анаэробное дыхание
Анаэробное дыхание — биохимический процесс окисления органических субстратов или молекулярного водорода с использованием в дыхательной ЭТЦ в качестве конечного акцептора электронов вместо O2 других окислителей неорганической или органической природы. Как и в случае аэробного дыхания, выделяющаяся в ходе реакции свободная энергия запасается в виде трансмембранного протонного потенциала, использующегося АТФ-синтазой для синтеза АТФ.
Осуществляется прокариотами (в редких случаях — и эукариотами) в анаэробных условиях. При этом факультативные анаэробы используют акцепторы электронов с высоким окислительно-восстановительным потенциалом (NO3 − , NO2 − , Fe 3+ , фумарат, диметилсульфоксид и т. д.), у них это дыхание конкурирует с энергетически более выгодным аэробным и подавляется кислородом. Акцепторы с низким окислительно-восстановительным потенциалом (сера, SO4 2− , CO2) применяются только строгими анаэробами, гибнущими при появлении в среде кислорода. В корневых системах многих растений при гипоксии и аноксии, вызванных затоплением посевов в результате длительных дождей или весенних паводков, развивается анаэробное дыхание с использованием в качестве акцепторов электронов альтернативных кислороду соединений, например нитратов. Установлено, что растения, произрастающие на полях, удобренных нитратными соединениями, переносят переувлажнение почвы и сопутствующую ему гипоксию лучше, нежели такие же растения без нитратной подкормки.
Механизмы окисления органических субстратов при анаэробном дыхании, как правило, аналогичны механизмам окисления при аэробном дыхании. Исключением является использование в качестве исходного субстрата ароматических соединений. Обычные пути их катаболизма требуют молекулярного кислорода уже на первых стадиях, в анаэробных условиях осуществляются иные процессы, например, восстановительная деароматизация бензоил-КоА у Thauera aromatica с затратой энергии АТФ. Некоторые субстраты (например, лигнин) при анаэробном дыхании не могут использоваться.
Содержание
Нитратное и нитритное дыхание
Прокариоты обладают возможностью использовать в качестве акцептора электрона в дыхательной электронтранспортной цепи (ЭТЦ) вместо кислорода различные окисленные соединения азота. Ферментом, катализирующим финальную стадию транспорта электрона — его перенос на нитрат-анион — является нитратредуктаза. При использовании нитритов ферментов и путей его восстановления два:
- NO-образующая нитритредуктаза восстанавливает нитрит до оксида азота (II). Это одна из стадий денитрификации.
- диссимиляционного восстановления нитратов в аммоний или, как его называют в иностранной литературе, диссимиляционной или дыхательной аммонификации (в отечественной литературой аммонификацией называется процесс высвобождения аммиака из состава органических соединений, например, белков). Надо отметить, что ассимиляционное восстановление нитратов в аммоний (ассимиляционная нитратредукция или просто ассимиляция) — процесс включения нитратов после восстановления до аммония в состав органических веществ, широко распространённый у прокариот и некоторых групп эукариот (грибы, растения) — не сопряжён с получением энергии.
НАД·H, образовавшийся при гликолизе, в ЦТК или по иным механизмам и поступающий в дыхательную ЭТЦ, окисляется обычно НАД·H:убихинон-оксидоредуктазой, являющейся протонной помпой. Терминальные оксидоредуктазы, переносящие электрон на конечный акцептор, в отличие от цитохромоксидазы аэробной ЭТЦ, обычно не являются протонной помпой. Однако при переносе нитратредуктазой электрона с убихинона (или у ряда видов менахинона) на нитрат-анион происходит выделение двух протонов в периплазму (с убихинона) и связываение двух протонов в воду в цитоплазме. Таким образом создаётся дополнительный протонный градиент.
Аналогичным образом, связывая протоны в цитоплазме, создаёт градиент электрохимического потенциала нитритредуктаза. В то же время NO-редуктаза связывает протоны из периплазмы и её работа не сопряжена с образованием градиента потенциала [1] .
Больше путей переноса протонов через мембрану анаэробная ЭТЦ не содержит (в аэробной же их 3), в связи с чем нитратное дыхание по эффективности в расчёте на 1 моль глюкозы составляет лишь 70 % от аэробного. При поступлении в среду молекулярного кислорода бактерии переключаются на обычное дыхание.
Нитратное дыхание встречается, хотя и редко, и среди эукариот. Так, нитратное дыхание, сопровождающееся денитрификацией и выделением молекулярного азота, недавно открыто у фораминифер [2] . До этого нитратное дыхание с образованием N2O было описано у грибов Fusariumи Cylindrocarpon (см. [3] .
Сульфатное дыхание
В настоящее время известен ряд бактерий, способных окислять органические соединения или молекулярный водород в анаэробных условиях, используя в качестве акцепторов электронов в дыхательной цепи сульфаты, тиосульфаты, сульфиты, молекулярную серу. Этот процесс получил название диссимиляционной сульфатредукции, а бактерии, осуществляющие этот процесс — сульфатвосстанавливающих или сульфатредуцирующих.
Все сульфатвосстанавливающие бактерии — облигатные анаэробы.
Сульфатвосстанавливающие бактерии получают энергию в процессе сульфатного дыхания при переносе электронов в электронтранспортной цепи. Перенос электронов от окисляемого субстрата по электронтранспортной цепи сопровождается возникновением электрохимического градиента ионов водорода с последующим синтезом АТФ.
Подавляющее большинство бактерий этой группы хемоорганогетеротрофы. Источником углерода и донором электронов для них являются простые органические вещества — пируват, лактат, сукцинат, малат, а также некоторые спирты. У некоторых сульфатвосстанавливающих бактерий обнаружена способность к хемолитоавтотрофии, когда окисляемым субстратом является молекулярный водород.
Сульфатвосстанавливающие эубактерии широко распространены в анаэробных зонах водоёмов разного типа, в иле, в почвах, в пищеварительном тракте животных. Наиболее интенсивно восстановление сульфатов происходит в соленых озерах и морских лиманах, где почти нет циркуляции воды, и содержится много сульфатов. Сульфатвосстанавливающим эубактериям принадлежит ведущая роль в образовании сероводорода в природе и в отложении сульфидных минералов. Накопление в среде H2S часто приводит к отрицательным последствиям — в водоемах к гибели рыбы, в почвах к угнетению растений. С активностью сульфатвосстанавливающих эубактерий связана также коррозия в анаэробных условиях различного металлического оборудования, например, металлических труб.
Фумаратное дыхание
В качестве акцептора электронов может использоваться фумарат. Фумаратредуктаза сходна с нитритредуктазой: лишь вместо молибдоптерин содержащей субъединицы в её состав входит ФАД и гистидин содержащая субъединица. Трансмембранный протонный потенциал образуется аналогичным образом: перенос протонов не происходит, однако фумаратредуктаза связывает протоны в цитоплазме, а дегидрогеназы в начале ЭТЦ выделяют протоны в периплазму. Перенос электронов с дегидрогеназ на фумаратредуктазу происходит обычно через мембранный пул менохинонов.
Фумарат, как правило, отсутствует в природных местообитаниях и образуется самими микроорганизмами из аспартата, аспарагина, сахаров, малата и цитрата. В виду этого большинство бактерий, способных к фумаратному дыханию содержат фумаразу, аспартат:аммиак-лиазу и аспарагиназу, синтез которого контролирует чувствительный к молекулярному кислороду белок Fnr.
Фумаратное дыхание достаточно широко распространено среди эукариот, в частности у животных (среди животных, у которых оно описано — пескожил, мидии, аскарида, печеночная двуустка и др [3] .)
Железистое дыхание

Дыхание ацетогенных бактерий
Строго анаэробные ацетогенные бактерии родов Acetobacterium, Clostridium, Peptostreptococcus и др. способны получать энергию, окисляя водород углекислым газом. При этом две молекулы CO2 образуют ацетат. Энергия при этом запасается в виде трансмембранного градиента протонов (Clostridium sp.) или ионов натрия (Acetobacterium woodi). Для перевода его в энергию связей АТФ используется обычная H-транспортирующая АТФ-синтаза или Na-зависимая АТФ-синтаза соответственно.
Анаэробное дыхание у растений
Анаэробное дыхание, в частности нитратное, активируется в корневых системах некоторых растений в условиях аноксии и гипоксии. Однако если у многих бактерий и некоторых протистов и животных оно может быть основным и достаточным для получения энергии процессом (часто наряду с гликолизом), то у растений функционирует почти исключительно в стрессовых условиях. Так или иначе, но на полях, где в качестве удобрений были внесены нитраты, растения лучше переносят гипоксию, вызванную переувлажнением почвы из-за продолжительных дождей [4] [5] .
Анаэробное дыхание у грибов, протистов и животных
Среди животных анаэробное фумаратное дыхание встречается у некоторых паразитических и свободноживущих червей, ракообразных, моллюсков; нитратное дыхание известно среди грибов (например, у Fusarium) [1] [6] и свободноживущих протистов — например, фораминифер [2] [7] . Способность к анаэробному дыханию присутствует и у черепах, однако непродолжительное время
Источник



